
Статья 704 ГК РФ. Выполнение работы иждивением подрядчика
Новая редакция Ст. 704 ГК РФ
1. Если иное не предусмотрено договором подряда, работа выполняется иждивением подрядчика — из его материалов, его силами и средствами.
2. Подрядчик несет ответственность за ненадлежащее качество предоставленных им материалов и оборудования, а также за предоставление материалов и оборудования, обремененных правами третьих лиц.
Комментарий к Ст. 704 ГК РФ
1. Нормы комментируемой статьи, раскрывающие выполнение работ (по общему правилу) иждивением подрядчика, касаются только отношений между заказчиком и подрядчиком.
Термин «иждивение» в комментируемой статье использован в одном из его устаревших значений, которое, однако, традиционно для российского законодательства.
Наука.
Подряд выполняется за счет и риск подрядчика, или, как выражается закон, его «иждивением».
Г.Ф.Шершеневич
2. Пункт 1 комментируемой статьи непосредственно посвящен раскрытию такой составляющей выполнения работ иждивением подрядчика, как выполнение работ «за счет» подрядчика.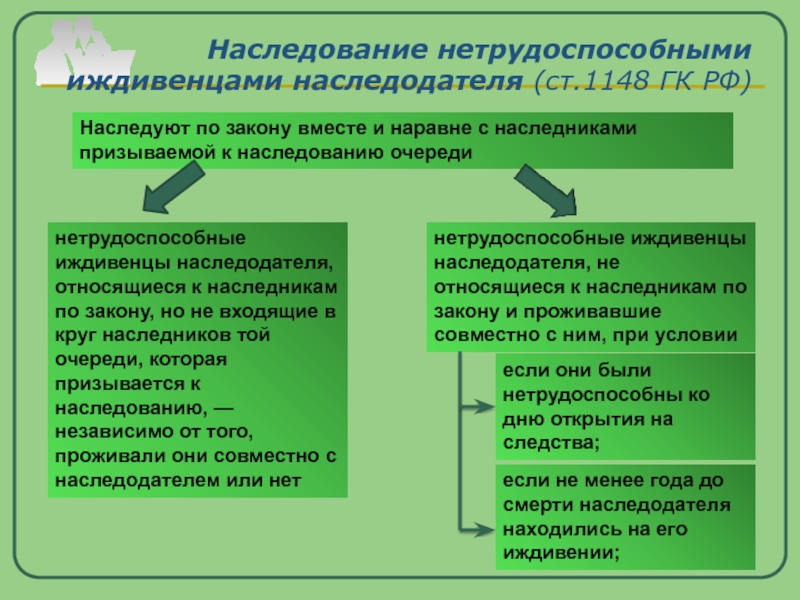
Правило о выполнении работ из материалов подрядчика означает, что подрядчик использует материалы, уже имеющиеся у него, или самостоятельно приобретает материалы от своего имени (о предоставлении материалов заказчиком — см. ст. ст. 713, 714 ГК РФ).
Требование к выполнению работ подрядчиком своими силами ни в коем случае не означает, что подрядчик обязан исполнить работы лично (напротив, в виде общего подхода установлено иное правило — см. п. 1 ст. 706 ГК РФ). Выполнение работ своими силами означает, что подрядчик сам организует рабочий процесс: добывает материалы, использует собственное или полученное в пользование оборудование, нанимает рабочих и т.п.
Требование к выполнению работ подрядчиком своими средствами касается прежде всего порядка оплаты работ: по общему правилу заказчик не финансирует работы, а оплачивает лишь их готовый результат (ст. 711 ГК РФ). Данное правило не означает, что подрядчик может использовать только собственные средства — он может получить кредитование со стороны третьих лиц.
3. Выполнение работ подрядчиком «за свой риск» как составляющей выполнения работ иждивением подрядчика непосредственно в нормах комментируемой статьи не раскрыт. Однако для подрядчика выполнение работ «за свой риск» неизбежно вытекает из выполнения им работ «за свой счет».
В частности, несение риска подрядчиком отражено в п. 2 комментируемой статьи о его ответственности за предоставленные материалы и оборудование (об ответственности подрядчика за качество предоставленных им материалов см. п. 5 ст. 723 ГК РФ). Несение риска подрядчиком находит свое отражение и в том, что оплате подлежит лишь результат его работы (см. ст. 711 ГК РФ). Самостоятельность подрядчика как проявление его общего риска отражена в п. 3 ст. 703 ГК.
Другой комментарий к Ст. 704 Гражданского кодекса Российской Федерации
1. В п. 1 настоящей статьи закреплено правило, известное ранее из ГК 1922 и 1964 гг., — подрядчик выполняет работу своим иждивением. В отличие от ГК 1964 г., в ст. 353 которого под иждивением подрядчика подразумевалось выполнение работ из его материалов и его средствами, ст.
2. Как и в предыдущих Гражданских кодексах, норма п. 1 сформулирована диспозитивно. Это означает, что если в договоре не предусмотрено, кто должен предоставлять материалы и оборудование, в процессе исполнения обязательства подрядчик не вправе требовать их от заказчика. Так, в одном из арбитражных дел подрядчик, нарушивший срок выполнения работ, для освобождения себя от ответственности ссылался на виновное поведение заказчика, выразившееся в непредставлении подрядчику материалов, машин, оборудования, необходимых для выполнения подрядных работ. Эти доводы ответчика не были приняты судом во внимание, так как при заключении договора стороны не пришли к единому согласию о том, кто обязан обеспечивать объект строительными и другими материалами.
3. Из п. 1 данной статьи вытекает правило, предусмотренное в п. 2, об ответственности подрядчика за ненадлежащее качество предоставленных им материалов и оборудования. При этом за качество предоставленного материала он отвечает по правилам об ответственности продавца за товары ненадлежащего качества, т.е. по ст. 475 ГК (п. 5 ст. 723 ГК РФ). (См. п. 5 комментария к ст. 723 ГК.)
4. Подрядчик отвечает не только за качество предоставленного материала и оборудования, но и за обременение их правами третьих лиц. Ими могут быть арендодатель, залогодержатель и т.д. Если они заявляют свои требования на материал или оборудование подрядчика, в связи с чем работа не может быть выполнена или заканчивается с нарушением срока, подрядчик несет ответственность перед заказчиком, например, в форме возмещения убытков.
Ст. 704 ГК РФ с Комментариями 2020-2021 года (новая редакция с последними изменениями)
1. Если иное не предусмотрено договором подряда, работа выполняется иждивением подрядчика — из его материалов, его силами и средствами.
2. Подрядчик несет ответственность за ненадлежащее качество предоставленных им материалов и оборудования, а также за предоставление материалов и оборудования, обремененных правами третьих лиц.
Комментарий к Ст. 704 ГК РФ
1. Согласно п. 1 комментируемой статьи подрядчик выполняет работу, если иное не предусмотрено договором подряда, собственным иждивением, т.е. из своих материалов, своими силами и средствами . Осуществляя работу собственными средствами, подрядчик не только несет необходимые затраты, связанные с выполнением работ, но и использует собственное оборудование, как это следует из п. 2 данной статьи.
———————————
Термин «собственное иждивение» в п. 1 комментируемой статьи применяется в традиционном его понимании.
Бесплатная юридическая консультация по телефонам:
Норма, содержащаяся в п. 1 комментируемой статьи, является диспозитивной, поэтому в договоре подряда стороны могут предусмотреть и иные варианты распределения прав и обязанностей. Так, согласно ст. 706 ГК РФ, если из закона или договора подряда не вытекает обязанность подрядчика выполнить предусмотренную в договоре работу лично, подрядчик вправе привлечь к исполнению своих обязательств других лиц (субподрядчиков), а в соответствии со ст. 707 ГК РФ на стороне подрядчика могут выступать одновременно два или более лиц. Согласно ст. 713 ГК РФ в случае предоставления заказчиком материала, необходимого для выполнения работы, подрядчик обязан использовать его экономно и расчетливо, после окончания работы представить заказчику отчет об израсходовании материала, а также возвратить его остаток либо с согласия заказчика уменьшить цену работы с учетом стоимости остающегося у подрядчика неиспользованного материала.
2. Поскольку по общему правилу выполнение работы осуществляется иждивением подрядчика, он несет ответственность за ненадлежащее качество предоставленных им материалов и оборудования (п. 2 комментируемой статьи). Согласно п. 5 ст. 723 ГК РФ подрядчик, предоставивший материал для выполнения работы, отвечает за его качество по правилам об ответственности продавца за товары ненадлежащего качества. Следовательно, заказчик вправе воспользоваться любой из возможностей, предоставленных покупателю ст. 475 ГК РФ.
Что касается правовых последствий ненадлежащего качества предоставленного подрядчиком оборудования, то они могут различаться в зависимости от наступивших ввиду использования некачественного оборудования результатов. Так, если использование некачественного оборудования повлекло ненадлежащее качество работы (недостатки), то к подрядчику могут применяться правила ст. 723 ГК РФ. В том случае, когда использование некачественного оборудования повлекло нарушение сроков выполнения работ, к подрядчику могут применяться меры воздействия, предусмотренные ст. 708 ГК РФ, а если такое использование привело к тому, что подрядчик не приступает своевременно к исполнению договора подряда или выполняет работу настолько медленно, что окончание ее к сроку становится явно невозможным, заказчик вправе отказаться от исполнения договора и потребовать возмещения убытков (п. 3 ст. 715 ГК).
723 ГК РФ. В том случае, когда использование некачественного оборудования повлекло нарушение сроков выполнения работ, к подрядчику могут применяться меры воздействия, предусмотренные ст. 708 ГК РФ, а если такое использование привело к тому, что подрядчик не приступает своевременно к исполнению договора подряда или выполняет работу настолько медленно, что окончание ее к сроку становится явно невозможным, заказчик вправе отказаться от исполнения договора и потребовать возмещения убытков (п. 3 ст. 715 ГК).
Вместе с тем нет достаточных оснований для того, чтобы распространять положения об ответственности подрядчика за ненадлежащее качество предоставленных им материалов и оборудования, приведенные в п. 2 комментируемой статьи, на его собственные силы и средства, во всяком случае на ту часть средств, которая включает необходимые затраты, осуществляемые на выполнение работ. Дело в том, что использование не имеющего соответствующей квалификации персонала либо неосуществление необходимых затрат для надлежащего выполнения работ подрядчиком может привести к самым разным правовым последствиям в зависимости от наступивших в каждом конкретном случае результатов.
3. С выполнением по общему правилу работы иждивением подрядчика связывается также его ответственность за предоставление материалов и оборудования, обремененных правами третьих лиц. Под материалами и оборудованием, обремененными правами третьих лиц, понимается наличие на них различных вещных или обязательственных прав (право собственности, право залога и т.д.) у лиц, которые не являются сторонами договора подряда. Указанные права могут препятствовать надлежащему исполнению договора подрядчиком, поэтому, как и в случаях с оборудованием ненадлежащего качества, а также с ненадлежащим использованием его собственных сил и средств, правовые последствия здесь тоже будут различаться в зависимости от наступивших в каждом конкретном случае результатов. В связи с этим представляется, что указание в п. 2 комментируемой статьи на то, что подрядчик несет ответственность за ненадлежащее качество предоставленных им материалов и оборудования, а также за предоставление материалов и оборудования, обремененных правами третьих лиц, служит распределению между сторонами договора подряда рисков неисполнения или ненадлежащего исполнения обязательств. То есть риск неблагоприятных последствий в случае неисполнения или ненадлежащего исполнения обязательств по договору подряда по причинам ненадлежащего качества предоставленных подрядчиком материалов и оборудования, а также в случае предоставления им материалов и оборудования, обремененных правами третьих лиц, всегда возлагается на подрядчика.
То есть риск неблагоприятных последствий в случае неисполнения или ненадлежащего исполнения обязательств по договору подряда по причинам ненадлежащего качества предоставленных подрядчиком материалов и оборудования, а также в случае предоставления им материалов и оборудования, обремененных правами третьих лиц, всегда возлагается на подрядчика.
1. По договору пожизненного содержания с иждивением получатель ренты — гражданин передает принадлежащие ему жилой дом, квартиру, земельный участок или иную недвижимость в собственность плательщика ренты, который обязуется осуществлять пожизненное содержание с иждивением гражданина и (или) указанного им третьего лица (лиц). 2. К договору пожизненного содержания с иждивением применяются правила о пожизненной ренте, если иное не предусмотрено правилами настоящего параграфа. Судебная практика по статье 601 ГК РФ.
|
 по делу № 2-588/2020
по делу № 2-588/2020  ..
.. В соответствии со ст. 601 ГК РФ по договору пожизненного содержания с иждивением получатель ренты — гражданин передает принадлежащие ему жилой дом, квартиру, земельный участок или иную недвижимость в собственность плательщика ренты, который …
В соответствии со ст. 601 ГК РФ по договору пожизненного содержания с иждивением получатель ренты — гражданин передает принадлежащие ему жилой дом, квартиру, земельный участок или иную недвижимость в собственность плательщика ренты, который … по делу № 2-309/2020
по делу № 2-309/2020  С. удовлетворить.
Расторгнуть договор пожизненного …
С. удовлетворить.
Расторгнуть договор пожизненного … На основании ст. 601 ГК РФ по договору пожизненного содержания с иждивением получатель ренты — гражданин передает принадлежащие ему жилой дом, квартиру, земельный участок или иную недвижимость в собственность плательщика ренты, который …
На основании ст. 601 ГК РФ по договору пожизненного содержания с иждивением получатель ренты — гражданин передает принадлежащие ему жилой дом, квартиру, земельный участок или иную недвижимость в собственность плательщика ренты, который …Статья 602. Обязанность по предоставлению содержания с иждивением
1. Обязанность плательщика ренты по предоставлению содержания с иждивением может включать обеспечение потребностей в жилище, питании и одежде, а если этого требует состояние здоровья гражданина, также и уход за ним. Договором пожизненного содержания с иждивением может быть также предусмотрена оплата плательщиком ренты ритуальных услуг.
Обязанность плательщика ренты по предоставлению содержания с иждивением может включать обеспечение потребностей в жилище, питании и одежде, а если этого требует состояние здоровья гражданина, также и уход за ним. Договором пожизненного содержания с иждивением может быть также предусмотрена оплата плательщиком ренты ритуальных услуг.
2. В договоре пожизненного содержания с иждивением должна быть определена стоимость всего объема содержания с иждивением. При этом стоимость общего объема содержания в месяц по договору пожизненного содержания с иждивением, предусматривающему отчуждение имущества бесплатно, не может быть менее двух установленных в соответствии с законом величин прожиточного минимума на душу населения в соответствующем субъекте Российской Федерации по месту нахождения имущества, являющегося предметом договора пожизненного содержания с иждивением, а при отсутствии в соответствующем субъекте Российской Федерации указанной величины не менее двух установленных в соответствии с законом величин прожиточного минимума на душу населения в целом по Российской Федерации.
3. При разрешении спора между сторонами об объеме содержания, которое предоставляется или должно предоставляться гражданину, суд должен руководствоваться принципами добросовестности и разумности.
Комментарий к ст. 602 ГК РФ
1. Примерный перечень обязанностей плательщика ренты по предоставлению содержания приводится в п. 1 коммент. ст. Конкретным договором данный перечень может быть расширен, сокращен или уточнен применительно к удовлетворению каждой из потребностей получателя ренты.
2. Пункт 2 ст. 602 требует, чтобы в договоре была указана стоимость всего объема содержания с иждивением, которая не может быть менее двух минимальных размеров оплаты труда. Данное правило императивно и установлено в интересах получателей ренты. Общая стоимость содержания подлежит индексации в соответствии с ростом минимального размера оплаты труда.
По смыслу закона стороны могут в договоре не только конкретизировать условия, качество, форму и т. п. каждого вида предоставления, но и зафиксировать в договоре их величину в стоимостном выражении. Вместе с тем характер данного договора предполагает, что виды и объем содержания, даже если они четко оговорены договором, не могут рассматриваться в качестве неизменных величин. Напротив, они могут меняться в зависимости от изменения разумных и обоснованных потребностей получателей ренты.
п. каждого вида предоставления, но и зафиксировать в договоре их величину в стоимостном выражении. Вместе с тем характер данного договора предполагает, что виды и объем содержания, даже если они четко оговорены договором, не могут рассматриваться в качестве неизменных величин. Напротив, они могут меняться в зависимости от изменения разумных и обоснованных потребностей получателей ренты.
3. Периодичность предоставления содержания законом особо не оговаривается, так как данный вопрос решается исходя из необходимости обеспечения нормальных естественных потребностей человека. Вместе с тем стороны могут регламентировать и этот аспект своих взаимоотношений. Что касается стоимости объема содержания с иждивением, то она рассчитывается, по общему правилу, помесячно, если сторонами не предусмотрено иное.
4. Пункт 3 ст. 602 предоставляет возможность разрешения спора об объеме содержания судом. Это подтверждает сделанный ранее вывод о том, что объем содержания может меняться в связи, напр. , с изменением состояния здоровья иждивенца, необходимостью ухода за ним, под влиянием внешних обстоятельств и т.п. Разрешая данный спор, суд должен учесть многие обстоятельства, включая стоимость переданного имущества, факт передачи имущества за плату или бесплатно, материальное положение иждивенца и плательщика ренты и т.д. Но, в конечном счете, руководствуясь принципами добросовестности и разумности, суд должен исходить из основной цели договора, состоящей в обеспечении иждивенцу хотя бы сносных условий существования.
, с изменением состояния здоровья иждивенца, необходимостью ухода за ним, под влиянием внешних обстоятельств и т.п. Разрешая данный спор, суд должен учесть многие обстоятельства, включая стоимость переданного имущества, факт передачи имущества за плату или бесплатно, материальное положение иждивенца и плательщика ренты и т.д. Но, в конечном счете, руководствуясь принципами добросовестности и разумности, суд должен исходить из основной цели договора, состоящей в обеспечении иждивенцу хотя бы сносных условий существования.
Судебная практика по статье 602 ГК РФ
Определение Судебной коллегии по гражданским делам Верховного Суда Российской Федерации от 28.01.2020 N 18-КГ19-140, 2-4483/2018Разрешая спор и удовлетворяя исковые требования Феоктистовой В. М., суд первой инстанции, руководствуясь положениями статей 601, 602 и 605 Гражданского кодекса Российской Федерации, сославшись на акт осмотра спорного домовладения от 1 ноября 2017 г. (л.д. 175), а также на показания свидетелей Евтеевой Т.Н., Приходько Н.Н., Чубовой А.А., исходил из того, что ответчица не представила доказательств надлежащего выполнения обязанностей по содержанию истицы в натуре.
М., суд первой инстанции, руководствуясь положениями статей 601, 602 и 605 Гражданского кодекса Российской Федерации, сославшись на акт осмотра спорного домовладения от 1 ноября 2017 г. (л.д. 175), а также на показания свидетелей Евтеевой Т.Н., Приходько Н.Н., Чубовой А.А., исходил из того, что ответчица не представила доказательств надлежащего выполнения обязанностей по содержанию истицы в натуре.
Определение Конституционного Суда РФ от 28.06.2018 N 1524-О
Применение же пункта 2 статьи 602 ГК Российской Федерации в деле с участием заявителя представленными материалами не подтверждается.
Исходя из изложенного и руководствуясь пунктом 2 статьи 43, частью первой статьи 79, статьями 96 и 97 Федерального конституционного закона «О Конституционном Суде Российской Федерации», Конституционный Суд Российской Федерации
Договор пожизненного содержания с иждивением как разновидность договора ренты
Наследование, дарение и рента — три наиболее распространенных среди населения способа распоряжения своим имуществом. Договору ренты посвящена 33 глава Гражданского кодекса Российской Федерации.
Договору ренты посвящена 33 глава Гражданского кодекса Российской Федерации.
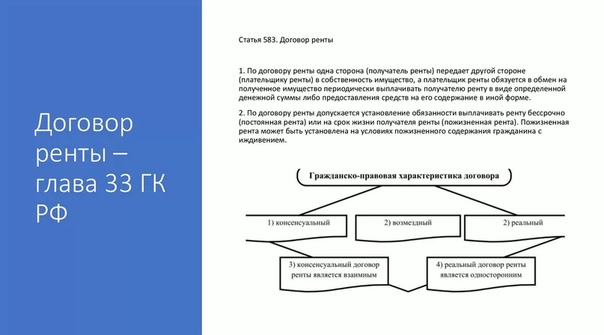
По договору ренты одна сторона (получатель ренты) передает другой стороне (плательщику ренты) в собственность имущество, а плательщик ренты обязуется в обмен на полученное имущество периодически выплачивать получателю ренту в виде определенной денежной суммы либо предоставления средств на его содержание в иной форме (п. 1 ст. 583 ГК РФ).
Договор ренты, как и договор купли-продажи, мены, дарения, предусматривает отчуждение имущества и по этому признаку сходен с указанными договорами. В случае, когда договором ренты предусматривается передача имущества за плату, к отношениям сторон по передаче и оплате применяются правила о купле-продаже (глава 30 ГК РФ), а в случае, когда такое имущество передается бесплатно, правила о договоре дарения (глава 32 ГК РФ) постольку, поскольку иное не установлено законодательством и не противоречит существу договора ренты. Можно сказать, что он относится к группе договоров по отчуждению имущества, но тем не менее является самостоятельным видом договора.
В Гражданском кодексе РФ выделены три разновидности рентного договора: 1) постоянная рента; 2) пожизненная рента; 3) пожизненное содержание с иждивением. Существенные условия всех трех видов ренты совпадают. Конечно, имеются и отличия в форме предоставления содержания, его минимальном размере, сроках предоставления, субъектном составе обязательств, возможностях правопреемства и выкупа ренты, последствиях риска случайной гибели имущества и т.д.
Существенно отличаются друг от друга договоры ренты, по которым под выплату ренты передано недвижимое и движимое имущество. Различия между ними проходят по способам оформления, по наличию или отсутствию признака следования ренты за имуществом при его отчуждении, по предусмотренным законом способам обеспечения исполнения обязательств плательщика ренты и т.д. Одно из вышеперечисленных отличий форма договора. Так договор ренты подлежит обязательному нотариальному удостоверению, а договор, предусматривающий отчуждение недвижимого имущества под выплату ренты, подлежит также государственной регистрации.
Одним из разновидностей договора ренты, подлежащим государственной регистрации, является договор пожизненного содержания с иждивением. По договору пожизненного содержания с иждивением получатель ренты — гражданин передает принадлежащие ему жилой дом, квартиру, земельный участок или иную недвижимость в собственность плательщика ренты, который обязуется осуществлять пожизненное содержание с иждивением гражданина и (или) указанного им третьего лица.
Особенность данного вида договора, так же состоит в том, что срок данного вида обязательств определен продолжительностью жизни рентополучателя, если иное не вытекает из условий договора. Предметом такой разновидности ренты, как пожизненное содержание с иждивением, является недвижимое имущество в виде жилого дома, квартиры, земельного участка. Субъектом — получателем ренты может быть не только лицо, которое заключило договор и передало имущество, а также и лицо, на которое указано в договоре пожизненного содержания с иждивением. Обязанность плательщика ренты по предоставлению содержания с иждивением может включать обеспечение потребностей в жилище, питании и одежде, а если этого требует состояние здоровья гражданина, также и уход за ним. Договором пожизненного содержания с иждивением может быть также предусмотрена оплата плательщиком ренты ритуальных услуг. Условиями договора можно определить возможность замены предоставления содержания с иждивением в натуре выплатой в течение жизни гражданина периодических платежей в деньгах.
Обязанность плательщика ренты по предоставлению содержания с иждивением может включать обеспечение потребностей в жилище, питании и одежде, а если этого требует состояние здоровья гражданина, также и уход за ним. Договором пожизненного содержания с иждивением может быть также предусмотрена оплата плательщиком ренты ритуальных услуг. Условиями договора можно определить возможность замены предоставления содержания с иждивением в натуре выплатой в течение жизни гражданина периодических платежей в деньгах.
Поскольку данный вид договора является, в некотором роде доверительным, то законодатель, для защиты интересов рентополучателя, предусмотрел минимальный размер содержания в месяц. В договоре пожизненного содержания с иждивением должна быть определена стоимость всего объема содержания с иждивением. При этом стоимость общего объема содержания в месяц не может быть менее двух минимальных размеров оплаты труда, установленных законом. Также плательщик ренты вправе отчуждать, сдавать в залог или иным способом обременять недвижимое имущество, переданное ему в обеспечение пожизненного содержания, только с предварительного согласия получателя ренты.
На плательщика ренты законодатель также возложил обязанность принимать необходимые меры для того, чтобы в период предоставления пожизненного содержания с иждивением использование указанного имущества не приводило к снижению стоимости этого имущества.
Законодательством предусмотрено два случая прекращения обязательства пожизненного содержания с иждивением:
В первом случае обязательства прекращаются смертью получателя ренты.
Во втором случае обязательства прекращаются по решению сторон. Если одна из сторон, а именно, плательщик ренты, существенно нарушала свои обязательства, получатель ренты вправе потребовать возврата недвижимого имущества, переданного в обеспечение пожизненного содержания, либо выплаты ему выкупной цены. При этом плательщик ренты не вправе требовать компенсацию расходов, понесенных в связи с содержанием получателя ренты.
#Росреестр#РосреестрКалининград#договор ренты
Что такое договор пожизненного содержания с иждивением
09. 08.2020 15:02:00
08.2020 15:02:00
Что такое договор пожизненного содержания с иждивением
Статьи Гражданского кодекса, регулирующие этот вопрос, разъясняет старший помощник Новомосковского горпрокурора Анна Строкова
Согласно статье 601 Гражданского кодекса Российской Федерации (далее ГК РФ), по договору пожизненного содержания с иждивением получатель ренты — гражданин передает принадлежащие ему жилой дом, квартиру, земельный участок или иную недвижимость в собственность плательщика ренты, который обязуется осуществлять пожизненное содержание с иждивением гражданина и (или) указанного им третьего лица (лиц).
Как установлено статьей 602 ГК РФ, обязанность плательщика ренты по предоставлению содержания с иждивением может включать обеспечение потребностей в жилище, питании и одежде, а если этого требует состояние здоровья гражданина, также и уход за ним. Договором пожизненного содержания с иждивением может быть также предусмотрена оплата плательщиком ренты ритуальных услуг.
В договоре пожизненного содержания с иждивением должна быть определена стоимость всего объема содержания с иждивением.
Договором пожизненного содержания с иждивением может быть предусмотрена возможность замены предоставления содержания с иждивением в натуре выплатой в течение жизни гражданина периодических платежей в деньгах.
Плательщик ренты вправе отчуждать, сдавать в залог или иным способом обременять недвижимое имущество, переданное ему в обеспечение пожизненного содержания, только с предварительного согласия получателя ренты.
Плательщик ренты обязан принимать необходимые меры для того, чтобы в период предоставления пожизненного содержания с иждивением использование указанного имущества не приводило к снижению стоимости этого имущества.
Согласно ст. 605 ГК РФ, обязательство пожизненного содержания с иждивением прекращается смертью получателя ренты.
При существенном нарушении плательщиком ренты своих обязательств получатель ренты вправе потребовать возврата недвижимого имущества, переданного в обеспечение пожизненного содержания, либо выплаты ему выкупной цены на условиях, установленных статьей 594 настоящего Кодекса. При этом плательщик ренты не вправе требовать компенсацию расходов, понесенных в связи с содержанием получателя ренты.
При этом плательщик ренты не вправе требовать компенсацию расходов, понесенных в связи с содержанием получателя ренты.
Фото familymediationcenter.ca
Договор ренты и договор пожизненного содержания с иждивением
Договор ренты и договор пожизненного содержания с иждивением
Заключение договора ренты, как следствие приобретения в собственность имущества под выплату ренты, достаточно известный вид сделки на рынке жилой недвижимости.
Согласно п. 1 ст. 583 Гражданского кодекса Российской Федерации (ГК РФ) по договору ренты одна сторона (получатель ренты) передает другой стороне (плательщику ренты) в собственность имущество, а плательщик ренты обязуется в обмен на полученное имущество периодически выплачивать получателю ренты денежные средства в виде определенной суммы либо предоставлять средства на его содержание в иной форме.
Договор ренты является самостоятельным видом договора и носит материальный характер, так как помимо придания договору соответствующей формы для его заключения требуется передача имущества плательщику ренты. После передачи имущества получатель ренты не несет по договору никаких обязанностей, обладая лишь правами. Возмездность договора обусловлена тем, что имущество передается в обмен на предоставление содержания в виде определенной денежной суммы или в иной форме.
После передачи имущества получатель ренты не несет по договору никаких обязанностей, обладая лишь правами. Возмездность договора обусловлена тем, что имущество передается в обмен на предоставление содержания в виде определенной денежной суммы или в иной форме.
Договор ренты, безусловно, имеет свою специфику. Она выражается в особых правовых отношениях между участвующими в нем лицами, и эти отношения совершенно не свойственны другим договорным гражданско-правовым обязательствам. Их суть заключается в обязанности одного лица периодически предоставлять пожизненное или постоянное содержание другому лицу, для которого оно нередко является единственным или основным источником получения средств к существованию. Любая из сторон может получить встречное удовлетворение в меньшем размере, чем предоставлено ею. Считается, что, заключая рентный договор, стороны осознают данное обстоятельство и с ним заранее соглашаются. Поэтому противоречило бы самой сути рассматриваемого договора введение в него условия о том, что общий размер рентных платежей, который может быть выплачен получателю ренты, ограничивается стоимостью переданного имущества.
Под выплату ренты может быть передано и движимое имущество. В этой ситуации согласно п. 2 ст. 587 ГК РФ стороны обязаны в качестве существенного условия предусмотреть в договоре ренты какое-либо обеспечение исполнения плательщиком ренты своих обязанностей. Это может быть залог, неустойка, поручительство 3-го лица.
Форма договора ренты регулируется специальными правилами. Договор ренты должен быть нотариально удостоверен. Если под выплату ренты было передано недвижимое имущество, то согласно ст. 584 ГК РФ такой договор подлежит также и государственной регистрации. При осуществлении государственной регистрации договора ренты, на основе которого происходит переход прав на недвижимость, такому акту подлежат и сам факт перехода имущества в собственность плательщика ренты, и лежащее на этом имуществе обременение в виде обязанности выплачивать ренту или предоставлять содержание получателю ренты в иной форме. Статья 9 Федерального закона от 13 июля 2015 № 218-ФЗ «О государственной регистрации недвижимости» позволяет сделать вывод, что при заключении договора ренты производится запись о переходе права собственности на определенную недвижимость к новому правообладателю.
Если стороны нарушат требования о нотариальном удостоверении договора ренты, такой договор будет считаться недействительной сделкой. Даже если такой договор ренты будет нотариально удостоверен, но не зарегистрирован в установленном порядке, он будет считаться незаключенной сделкой.
Как правило, получателями ренты по договору обычно являются пожилые люди или иные малоимущие, социально незащищенные граждане. В связи с этим ст. 586 ГК РФ предусмотрено, что рента обременяет имущество, переданное под ее выплату. В случае отчуждения такого имущества плательщиком ренты его обязательства по договору ренты переходят на приобретателя имущества. Лицо, передавшее обремененное рентой недвижимое имущество в собственность другого лица, несет субсидиарную с ним ответственность по требованиям получателя ренты, возникшим в связи с нарушением договора ренты, если другим законом или договором не предусмотрена солидарная ответственность по этому обязательству.
Кроме того, согласно ст. 588 ГК РФ предусмотрена ответственность плательщика ренты в виде штрафных процентов в случае просрочки выплаты ренты плательщиком. Следует указать, что размер процентов может определяться самими сторонами в заключенном договоре ренты.
588 ГК РФ предусмотрена ответственность плательщика ренты в виде штрафных процентов в случае просрочки выплаты ренты плательщиком. Следует указать, что размер процентов может определяться самими сторонами в заключенном договоре ренты.
В главе 33 ГК РФ говорится о договоре ренты как обобщающем понятии, поскольку данный договор всегда существует в одной из трех разновидностей, а именно в виде постоянной ренты, пожизненной ренты или пожизненного содержания с иждивением.
На сегодняшний день практика заключения договоров ренты, а особенно договоров пожизненного содержания с иждивением, продолжает постепенно расширяться, что связанно в первую очередь с проблемами в экономической сфере жизни, ведь, как уже было сказано выше, получателями ренты, как правило, являются пожилые или малоимущие люди, и заключение данных договоров является для них единственным средством к существованию.
Пожизненное содержание с иждивением рассматривается как вид договора пожизненной ренты. Согласно договору получатель ренты (гражданин) передает принадлежащие ему жилой дом, квартиру, земельный участок или иную недвижимость в собственность плательщика ренты, который обязуется осуществлять пожизненное содержание с иждивением гражданина и (или) указанного им третьего лица (лиц) (п. 1 ст. 601 ГК РФ).
Согласно договору получатель ренты (гражданин) передает принадлежащие ему жилой дом, квартиру, земельный участок или иную недвижимость в собственность плательщика ренты, который обязуется осуществлять пожизненное содержание с иждивением гражданина и (или) указанного им третьего лица (лиц) (п. 1 ст. 601 ГК РФ).
Для договора пожизненного содержания с иждивением характерны следующие признаки вида ренты:
во-первых, в обеспечение пожизненного содержания с иждивением может отчуждаться лишь недвижимое имущество: жилой дом, квартира, земельный участок и т.п.;
во-вторых, рента предоставляется не в денежной форме, а в виде обеспечения повседневных жизненных потребностей получателя ренты. Договором может быть также предусмотрена оплата плательщиком ренты ритуальных услуг;
в-третьих, заключить договор пожизненного содержания с иждивением может лишь гражданин, которому принадлежит жилой дом, квартира, земельный участок или иная недвижимость.
Для данного вида договора ренты в качестве существенного условия необходимо указать стоимость всего объема содержания. Кроме того, законодательством установлен минимальный размер такого содержания: в месяц он не может быть менее двух минимальных размеров оплаты труда, установленных законом. В то же время законодательством разрешено сторонам предусмотреть в договоре возможность замены предоставления содержания с иждивением в натуре выплатой в течение жизни гражданина периодических платежей в деньгах.
Договор пожизненного содержания с иждивением, в отличие от других видов ренты, предполагает более тесную связь плательщика ренты и ее получателя. Если в других видах ренты отношения сторон ограничиваются периодической передачей (перечислением) денежных средств или иного оговоренного договором предоставления при отсутствии, как правило, личных контактов, то здесь стороны договора обычно постоянно взаимодействуют друг с другом. Возлагая на плательщика ренты обязанности по обеспечению потребностей иждивенца в жилище, питании, одежде и т. п., иждивенец, как правило, хорошо его знает и доверяет ему. Замена плательщика ренты на другое лицо может оказаться для иждивенца абсолютно неприемлемой. Поэтому любые действия плательщика ренты, которые могут повлечь за собой в конечном счете переход права собственности на имущество к другому лицу, должны быть согласно абз. 1 ст. 604 ГК РФ предварительно согласованы с получателем ренты.
п., иждивенец, как правило, хорошо его знает и доверяет ему. Замена плательщика ренты на другое лицо может оказаться для иждивенца абсолютно неприемлемой. Поэтому любые действия плательщика ренты, которые могут повлечь за собой в конечном счете переход права собственности на имущество к другому лицу, должны быть согласно абз. 1 ст. 604 ГК РФ предварительно согласованы с получателем ренты.
Если же имущество было отчуждено, сдано в залог или иным образом обременено без согласия иждивенца, он вправе требовать признания этих актов недействительными.
Срок действия обязательства пожизненного содержания определяется его природой и равен периоду жизни иждивенца.
При существенном нарушении плательщиком ренты своих обязательств по пожизненному содержанию с иждивением получателя ренты действует общее правило: плательщик ренты не имеет права требовать компенсации расходов, понесенных в связи с содержанием получателя ренты. Получатель ренты вправе потребовать возврата недвижимого имущества, переданного в обеспечение пожизненного содержания, либо выплаты ему выкупной цены на условиях, установленных ст. 594 ГК РФ.
594 ГК РФ.
Подводя итог, можно отметить существующие различия договора пожизненного содержания с иждивением от других видов договора ренты, а именно: под получение содержания иждивенец передает не любое имущество, как это возможно при постоянной или пожизненной ренте, а только недвижимое имущество. Передать имущество может один гражданин, а получателем ренты может выступить указанное им третье лицо – выгодоприобретатель. В договоре пожизненного содержания с иждивением форма предоставления содержания может быть в виде совершения действий по обеспечению потребностей в жилище, питании, одежде, уходе, если это требует состояние здоровья, оплата плательщиком ренты ритуальных услуг.
В заключение приведем несколько основных правил, которые позволили бы сторонам договора ренты и договора пожизненного содержания с иждивением уменьшить риск их правовых отношений: для плательщика ренты необходимо прежде всего в договоре составить четкий список обязательств, которые он будет нести по отношению к получателю ренты, и определить денежную сумму либо предоставление средств на содержание в иной форме. Заключая договор ренты или пожизненного содержания с иждивением, получатель ренты должен хорошо знать плательщика ренты и быть уверенным в честном и порядочном отношении, не допускающем обмана и мошеннических действий.
Заключая договор ренты или пожизненного содержания с иждивением, получатель ренты должен хорошо знать плательщика ренты и быть уверенным в честном и порядочном отношении, не допускающем обмана и мошеннических действий.
Наталья Максимчук,
главный специалист-эксперт
отдела регистрации недвижимости
Управления Росреестра
по Омской области,
государственный регистратор.
Зависимость GC в реальных и смоделированных данных. GC-зависимость в …
Рак — это заболевание, связанное с аберрантной пролиферацией клеток и ростом опухоли, возникающее в результате нарушения эпигенетически определенной, регулируемой и поддерживаемой идентичности клеток посредством генетических мутаций. Это основная причина смерти во всем мире, и большинство видов рака остаются неизлечимыми. Технологии Omics — это количественные аналитические анализы, которые позволяют проводить высококачественные и высокопроизводительные измерения различных аспектов клеточной регуляции, включая геномику, транскриптомику, эпигеномику, протеомику и метаболомику.Эти высокопроизводительные технологии изменили методы проведения исследований рака, что привело к огромным успехам в нашем понимании биологии рака и современных целевых методов лечения. Интегральный анализ наборов данных multi-omics в исследованиях рака требует использования специальных алгоритмов, инструментов анализа и визуализации. Они разрабатываются и применяются в междисциплинарных группах ученых и клиницистов, работающих над совместными проектами. Как технические сложности анализа данных и их интеграции, так и эффективное независимое изучение наблюдений всеми партнерами по проекту являются современными исследовательскими задачами.В данной диссертации представлены результаты, рассматривающие широкий спектр этих вопросов. Глава 1, Замена превосходства CNS-PNET четырьмя новыми молекулярно-определенными объектами, обусловленными структурными вариантами: примитивные нейроэктодермальные опухоли центральной нервной системы (CNS-PNET) были гетерогенным семейством детских опухолей головного мозга без гистопатологических маркеров, сложной диагностики и плохого прогноза.
Технологии Omics — это количественные аналитические анализы, которые позволяют проводить высококачественные и высокопроизводительные измерения различных аспектов клеточной регуляции, включая геномику, транскриптомику, эпигеномику, протеомику и метаболомику.Эти высокопроизводительные технологии изменили методы проведения исследований рака, что привело к огромным успехам в нашем понимании биологии рака и современных целевых методов лечения. Интегральный анализ наборов данных multi-omics в исследованиях рака требует использования специальных алгоритмов, инструментов анализа и визуализации. Они разрабатываются и применяются в междисциплинарных группах ученых и клиницистов, работающих над совместными проектами. Как технические сложности анализа данных и их интеграции, так и эффективное независимое изучение наблюдений всеми партнерами по проекту являются современными исследовательскими задачами.В данной диссертации представлены результаты, рассматривающие широкий спектр этих вопросов. Глава 1, Замена превосходства CNS-PNET четырьмя новыми молекулярно-определенными объектами, обусловленными структурными вариантами: примитивные нейроэктодермальные опухоли центральной нервной системы (CNS-PNET) были гетерогенным семейством детских опухолей головного мозга без гистопатологических маркеров, сложной диагностики и плохого прогноза. Моя работа в качестве вычислительного биолога способствовала всестороннему описанию этой сущности.В этом исследовании мы применили комплексный омический анализ данных метиломов, транскриптомов и геномов, показавший, что CNS-PNET представляют собой комбинацию большой группы неправильно диагностированных случаев от других субъектов и четырех новых молекулярно определенных объектов. Я показал, что эти новые сущности управляются различными и повторяющимися молекулярными драйверами, измененными различными механизмами структурных вариантов: онкогеном FOXR2 и опухолевыми супрессорами MN1, CIC и BCOR. Наши результаты способствовали исключению CNS-PNET как официально признанного онкологического объекта и признанию четырех новых детских опухолей головного мозга в классификации опухолей головного мозга Всемирной организации здравоохранения.Глава 2, SOPHIA, Обнаружение структурных перестроек на основе дополнительных сопоставлений и фоновой модели популяции: основываясь на моей работе по структурным вариациям в нашем исследовании CNS-PNETs, я разработал алгоритм SOPHIA для обнаружения SV в геномах рака на основе большого популяционного фона.
Моя работа в качестве вычислительного биолога способствовала всестороннему описанию этой сущности.В этом исследовании мы применили комплексный омический анализ данных метиломов, транскриптомов и геномов, показавший, что CNS-PNET представляют собой комбинацию большой группы неправильно диагностированных случаев от других субъектов и четырех новых молекулярно определенных объектов. Я показал, что эти новые сущности управляются различными и повторяющимися молекулярными драйверами, измененными различными механизмами структурных вариантов: онкогеном FOXR2 и опухолевыми супрессорами MN1, CIC и BCOR. Наши результаты способствовали исключению CNS-PNET как официально признанного онкологического объекта и признанию четырех новых детских опухолей головного мозга в классификации опухолей головного мозга Всемирной организации здравоохранения.Глава 2, SOPHIA, Обнаружение структурных перестроек на основе дополнительных сопоставлений и фоновой модели популяции: основываясь на моей работе по структурным вариациям в нашем исследовании CNS-PNETs, я разработал алгоритм SOPHIA для обнаружения SV в геномах рака на основе большого популяционного фона. написаны базы данных и соответствующий инструмент биоинформатики, позволяющий быстро обнаруживать SV из коротких наборов данных секвенирования генома рака. Позже SOPHIA стала стандартным инструментом для обнаружения структурных вариантов в рабочем процессе анализа генома рака DKFZ.Глава 3, EPISTEME, интерактивная и интегрирующая платформа для анализа, интерпретации и обмена данными мульти-омики: Во время разработки SOPHIA и моих исследований в проектах по анализу и интерпретации данных структурных вариантов я разработал опыт анализа данных структурных вариантов, обнаруженных SOPHIA, интегрируя они с различными слоями омики, такими как выражения генов, интерпретация, визуализация и обмен ими с соавторами, которые не были учеными-вычислителями. Основываясь на этом опыте и используя современные инструменты интерактивной визуализации данных, я разработал интерактивную платформу для комплексного анализа и визуализации данных омики под названием EPISTEME, с целью облегчить анализ данных омики учеными, обладающими концептуальными знаниями о онкологии, но не имеющими навыков программирования.
написаны базы данных и соответствующий инструмент биоинформатики, позволяющий быстро обнаруживать SV из коротких наборов данных секвенирования генома рака. Позже SOPHIA стала стандартным инструментом для обнаружения структурных вариантов в рабочем процессе анализа генома рака DKFZ.Глава 3, EPISTEME, интерактивная и интегрирующая платформа для анализа, интерпретации и обмена данными мульти-омики: Во время разработки SOPHIA и моих исследований в проектах по анализу и интерпретации данных структурных вариантов я разработал опыт анализа данных структурных вариантов, обнаруженных SOPHIA, интегрируя они с различными слоями омики, такими как выражения генов, интерпретация, визуализация и обмен ими с соавторами, которые не были учеными-вычислителями. Основываясь на этом опыте и используя современные инструменты интерактивной визуализации данных, я разработал интерактивную платформу для комплексного анализа и визуализации данных омики под названием EPISTEME, с целью облегчить анализ данных омики учеными, обладающими концептуальными знаниями о онкологии, но не имеющими навыков программирования. EPISTEME — это комплексный инструмент, объединяющий данные генома, транскриптома, метилома и протеома с клиническими метаданными в удобной веб-системе со статистическим анализом в браузере и выводом векторной графики публикационного качества. Глава 4, Интеграция SOPHIA-EPISTEME в проекты геномики рака DKFZ раскрывает новые подтипы заболеваний и информацию о разных типах рака: благодаря интеграции SOPHIA и EPISTEME в систему комплексного анализа данных омики моя работа выявила новые онкогены, активируемые захватом энхансера, и обнаружила новые молекулярные определены подтипы рефрактерной множественной миеломы (захват энхансера MYCN посредством перестройки иммуноглобулина в качестве замены MYC), острого миелоидного лейкоза у взрослых (активация MNX1 посредством захвата энхансера, предположительно действующего как механизм блока дифференциации) и детской нейробластомы (активация ATOh2 посредством предполагаемого захвата энхансера) Замена MYCN) в проектах, поддерживаемых DKFZ Гейдельбергским центром персонализированной онкологии (DKFZ-HIPO) и программами исследования рака Немецкого общества детской онкологии и гематологии (GPOH).
EPISTEME — это комплексный инструмент, объединяющий данные генома, транскриптома, метилома и протеома с клиническими метаданными в удобной веб-системе со статистическим анализом в браузере и выводом векторной графики публикационного качества. Глава 4, Интеграция SOPHIA-EPISTEME в проекты геномики рака DKFZ раскрывает новые подтипы заболеваний и информацию о разных типах рака: благодаря интеграции SOPHIA и EPISTEME в систему комплексного анализа данных омики моя работа выявила новые онкогены, активируемые захватом энхансера, и обнаружила новые молекулярные определены подтипы рефрактерной множественной миеломы (захват энхансера MYCN посредством перестройки иммуноглобулина в качестве замены MYC), острого миелоидного лейкоза у взрослых (активация MNX1 посредством захвата энхансера, предположительно действующего как механизм блока дифференциации) и детской нейробластомы (активация ATOh2 посредством предполагаемого захвата энхансера) Замена MYCN) в проектах, поддерживаемых DKFZ Гейдельбергским центром персонализированной онкологии (DKFZ-HIPO) и программами исследования рака Немецкого общества детской онкологии и гематологии (GPOH).
Сборка мусора и циклические ссылки в Java
1. Обзор
В этой быстрой статье мы увидим, как JVM обеспечивает сбор недостижимых, но циклических ссылок.
Сначала мы исследуем различные типы алгоритмов сборки мусора. После этого мы посмотрим, как циклические ссылки обрабатываются в JVM.
Также стоит упомянуть, что сборщик мусора не является частью спецификации JVM и оставлен на усмотрение разработчика. Следовательно, каждая реализация JVM может иметь разные стратегии сборки мусора или вообще не иметь.
В этой статье мы сосредоточимся на одной конкретной реализации JVM: JVM HotSpot. Мы также можем использовать термины JVM и HotSpot JVM как взаимозаменяемые на протяжении всей статьи.
2. Подсчет ссылок
Алгоритмы GC с подсчетом ссылок связывают счетчик ссылок с каждым объектом. Эти алгоритмы считают объект активным, пока количество ссылок на этот объект больше нуля . Обычно среда выполнения сохраняет счетчик ссылок в заголовке объекта.
Обычно среда выполнения сохраняет счетчик ссылок в заголовке объекта.
В очень наивной реализации каждая новая ссылка на объект должна запускать атомарное приращение счетчика ссылок. Точно так же каждое новое разыменование должно запускать атомарный декремент.
Язык программирования Swift использует форму подсчета ссылок для управления памятью. Кроме того, в JVM нет алгоритма GC, основанного на подсчете ссылок.
2.1. Плюсы и минусы
С другой стороны, подсчет ссылок может распределять затраты на управление памятью на протяжении жизненного цикла приложения, поскольку периодических сбоев сборщика мусора (почти) не возникает.Кроме того, он потенциально может уничтожить объекты, как только их счетчик ссылок достигнет нуля, и превратиться в мусор.
Подсчет ссылок — это тоже не бесплатный обед. В наивной реализации обновление счетчика ссылок может быть неэффективным, поскольку нам нужно увеличивать или уменьшать его атомарно. Некоторые оптимизации могут сделать подсчет ссылок более эффективным в этом отношении, например, подходы отложенного или буферизованного подсчета ссылок.
Некоторые оптимизации могут сделать подсчет ссылок более эффективным в этом отношении, например, подходы отложенного или буферизованного подсчета ссылок.
Однако есть еще одна серьезная проблема с подсчетом ссылок: он не может вернуть циклические ссылки .
Например, предположим, что объект A ссылается на объект B и наоборот. Даже если A и B станут недоступными для остальной части графа объектов, их счетчик ссылок никогда не достигнет нуля. Это потому, что они по-прежнему ссылаются друг на друга.
Как оказалось, такого рода циклические ссылки довольно распространены в информатике. Например, давайте рассмотрим следующий двусвязный список. Сначала другой объект имеет ссылку на список:
Связанный список доступен из объекта D, , поэтому его не следует собирать, а счетчики ссылок согласованы с этим ожиданием.Теперь предположим, что объект D сам становится недоступным:
Несмотря на то, что связанный список теперь также недоступен, количество ссылок для его компонентов больше одного. Следовательно, с этой наивной реализацией подсчета ссылок среда выполнения не будет рассматривать этот связанный список как мусор, даже если это .
Следовательно, с этой наивной реализацией подсчета ссылок среда выполнения не будет рассматривать этот связанный список как мусор, даже если это .
3. Трассировка GC
Сборщики трассировки определят доступность объектов, отслеживая их из набора корневых объектов, известных как корни GC .Если объект доступен из корневого объекта прямо или косвенно, он будет считаться живым. Остальные недоступны и кандидатов на взыскание:
Вот как работает простой сборщик трассировки. Начиная с корней сборщика мусора, он рекурсивно проходит по графу объектов, пока не останется больше серых объектов для посещения. В конце концов, он считает все белые объекты недоступными и кандидатами на сбор. Это простое описание алгоритма трехцветной маркировки.
Мы можем думать о корнях GC как об объектах, которые, как мы уверены, живы .Например, это некоторые корни GC в Java и JVM:
- Локальные переменные или что-то еще, к чему сейчас относятся кадры стека.
 Эти переменные используются выполняемыми в данный момент методами, поэтому мы не хотим их собирать
Эти переменные используются выполняемыми в данный момент методами, поэтому мы не хотим их собирать - Живые резьбы
- Статические переменные
- Классы, загруженные системным загрузчиком классов
- JNI, местные и глобальные
 Эти переменные используются выполняемыми в данный момент методами, поэтому мы не хотим их собирать
Эти переменные используются выполняемыми в данный момент методами, поэтому мы не хотим их собирать Сборщики трассировки, в отличие от сборщиков подсчета ссылок, будут периодически выполнять процесс сбора.Таким образом, в большинстве случаев распределения и назначения должны работать быстро. Однако, когда запускается сборщик мусора, может возникнуть некоторая икота.
С другой стороны, эти алгоритмы сборки мусора не пострадают от циклических ссылок . Вместо того, чтобы считать ссылки на каждый объект, они пересекают граф объектов, начиная с корней GC. Следовательно, даже если есть несколько циклических ссылок, объекты будут собираться до тех пор, пока они недоступны, как показано на диаграмме выше.
Интересно, что использование сборщика резервных копий трассировки в тандеме с GC подсчета ссылок является одним из традиционных подходов для исправления циклических ссылок при подсчете ссылок.
3.1. JVM HotSpot
Все реализации GC в JVM HotSpot на момент написания этой статьи являются сборщиками трассировки, включая CMS, G1 и ZGC . Таким образом, JVM не будет страдать от проблемы циклических ссылок. Это ключевой вывод из этой статьи!
4. Заключение
В этой быстрой статье мы увидели, как JVM обрабатывает циклические ссылки.
Для более подробной информации о сборке мусора настоятельно рекомендуется ознакомиться с руководством по сборке мусора.
android — «Превышен предел накладных расходов GC» после добавления зависимости
Проблема заключается в ограничении формата dex , а именно в 65536 ограничении на количество ссылок на методы .
Чтобы диагностировал проблему, увеличьте объем памяти до гораздо большего объема (включение отладки / подробного вывода не помогает, как ни странно). В моем случае я добавил в конфигурацию android-maven-plugin следующее:
-Xmx4096M
После выбора достаточно большого размера кучи сообщение изменится и будет содержать что-то вроде этого:
[ИНФОРМАЦИЯ] ошибка записи вывода: слишком много методов: 172296; макс - 65536. По пакету:
[ИНФОРМАЦИЯ] 20 android.app
[ИНФОРМАЦИЯ] 18 android.content
[ИНФОРМАЦИЯ] 1 android.content.pm
[ИНФОРМАЦИЯ] 7 android.content.res
[ИНФОРМАЦИЯ] 7 android.hardware
[ИНФОРМАЦИЯ] 38 android.media
[ИНФОРМАЦИЯ] 20 android.opengl
[ИНФОРМАЦИЯ] 11 android.os
[ИНФОРМАЦИЯ] 1 android.text.method
[ИНФОРМАЦИЯ] 8 android.util
[ИНФОРМАЦИЯ] 38 android.view
[ИНФОРМАЦИЯ] 2 android.view.inputmethod
[ИНФОРМАЦИЯ] 21 android.widget
...
[ИНФОРМАЦИЯ] 3 com.google.common.annotations
[ИНФОРМАЦИЯ] 746 com.google.common.base
[ИНФОРМАЦИЯ] 9 ком.google.common.base.internal
[ИНФОРМАЦИЯ] 833 com.google.common.cache
[ИНФОРМАЦИЯ] 8478 com.google.common.collect
[ИНФОРМАЦИЯ] 50 com.google.common.eventbus
[ИНФОРМАЦИЯ] 385 com.google.common.hash
[ИНФОРМАЦИЯ] 597 com.google.common.io
[ИНФОРМАЦИЯ] 92 com.google.common.math
[ИНФОРМАЦИЯ] 134 com.google.common.net
[ИНФОРМАЦИЯ] 521 com.google.common.primitives
[ИНФОРМАЦИЯ] 404 com.google.common.reflect
[ИНФОРМАЦИЯ] 954 com.google.common.util.concurrent
[ИНФОРМАЦИЯ] 16 по умолчанию
[ИНФОРМАЦИЯ] 227 it. unimi.dsi.fastutil
[ИНФОРМАЦИЯ] 1370 ит.уними.dsi.fastutil.booleans
[ИНФОРМАЦИЯ] 17249 it.unimi.dsi.fastutil.bytes
[ИНФОРМАЦИЯ] 17249 it.unimi.dsi.fastutil.chars
[ИНФОРМАЦИЯ] 17279 it.unimi.dsi.fastutil.doubles
[ИНФОРМАЦИЯ] 17263 it.unimi.dsi.fastutil.floats
[ИНФОРМАЦИЯ] 17309 it.unimi.dsi.fastutil.ints
[ИНФОРМАЦИЯ] 628 it.unimi.dsi.fastutil.io
[ИНФОРМАЦИЯ] 17315 it.unimi.dsi.fastutil.longs
[ИНФОРМАЦИЯ] 26514 it.unimi.dsi.fastutil.objects
[ИНФОРМАЦИЯ] 17257 it.unimi.dsi.fastutil.shorts
[ИНФОРМАЦИЯ] 2 java.awt
[ИНФОРМАЦИЯ] 5 java.awt.datatransfer
[ИНФОРМАЦИЯ] 206 java.io
[ИНФОРМАЦИЯ] 346 java.язык
[ИНФОРМАЦИЯ] 10 java.lang.ref
[ИНФОРМАЦИЯ] 71 java.lang.reflect
[ИНФОРМАЦИЯ] 27 java.math
[ИНФОРМАЦИЯ] 26 java.net
[ИНФОРМАЦИЯ] 108 java.nio
[ИНФОРМАЦИЯ] 6 java.nio.channels
[ИНФОРМАЦИЯ] 2 java.nio.charset
[ИНФОРМАЦИЯ] 13 java.security
[ИНФОРМАЦИЯ] 2 java.text
[ИНФОРМАЦИЯ] 382 java.util
[ИНФОРМАЦИЯ] 128 java.util.concurrent
[ИНФОРМАЦИЯ] 33 java.util.concurrent.atomic
[ИНФОРМАЦИЯ] 40 java.util.concurrent. locks
[ИНФОРМАЦИЯ] 6 java.util.jar
[ИНФОРМАЦИЯ] 6 java.util.logging
[ИНФОРМАЦИЯ] 10 java.util.regex
[ИНФОРМАЦИЯ] 20 java.util.zip
[ИНФОРМАЦИЯ] 26 javax.microedition.khronos.egl
[ИНФОРМАЦИЯ] 188 javax.microedition.khronos.opengles
[ИНФОРМАЦИЯ] 7 вс.misc
 По пакету:
[ИНФОРМАЦИЯ] 20 android.app
[ИНФОРМАЦИЯ] 18 android.content
[ИНФОРМАЦИЯ] 1 android.content.pm
[ИНФОРМАЦИЯ] 7 android.content.res
[ИНФОРМАЦИЯ] 7 android.hardware
[ИНФОРМАЦИЯ] 38 android.media
[ИНФОРМАЦИЯ] 20 android.opengl
[ИНФОРМАЦИЯ] 11 android.os
[ИНФОРМАЦИЯ] 1 android.text.method
[ИНФОРМАЦИЯ] 8 android.util
[ИНФОРМАЦИЯ] 38 android.view
[ИНФОРМАЦИЯ] 2 android.view.inputmethod
[ИНФОРМАЦИЯ] 21 android.widget
...
[ИНФОРМАЦИЯ] 3 com.google.common.annotations
[ИНФОРМАЦИЯ] 746 com.google.common.base
[ИНФОРМАЦИЯ] 9 ком.google.common.base.internal
[ИНФОРМАЦИЯ] 833 com.google.common.cache
[ИНФОРМАЦИЯ] 8478 com.google.common.collect
[ИНФОРМАЦИЯ] 50 com.google.common.eventbus
[ИНФОРМАЦИЯ] 385 com.google.common.hash
[ИНФОРМАЦИЯ] 597 com.google.common.io
[ИНФОРМАЦИЯ] 92 com.google.common.math
[ИНФОРМАЦИЯ] 134 com.google.common.net
[ИНФОРМАЦИЯ] 521 com.google.common.primitives
[ИНФОРМАЦИЯ] 404 com.google.common.reflect
[ИНФОРМАЦИЯ] 954 com.google.common.util.concurrent
[ИНФОРМАЦИЯ] 16 по умолчанию
[ИНФОРМАЦИЯ] 227 it.
По пакету:
[ИНФОРМАЦИЯ] 20 android.app
[ИНФОРМАЦИЯ] 18 android.content
[ИНФОРМАЦИЯ] 1 android.content.pm
[ИНФОРМАЦИЯ] 7 android.content.res
[ИНФОРМАЦИЯ] 7 android.hardware
[ИНФОРМАЦИЯ] 38 android.media
[ИНФОРМАЦИЯ] 20 android.opengl
[ИНФОРМАЦИЯ] 11 android.os
[ИНФОРМАЦИЯ] 1 android.text.method
[ИНФОРМАЦИЯ] 8 android.util
[ИНФОРМАЦИЯ] 38 android.view
[ИНФОРМАЦИЯ] 2 android.view.inputmethod
[ИНФОРМАЦИЯ] 21 android.widget
...
[ИНФОРМАЦИЯ] 3 com.google.common.annotations
[ИНФОРМАЦИЯ] 746 com.google.common.base
[ИНФОРМАЦИЯ] 9 ком.google.common.base.internal
[ИНФОРМАЦИЯ] 833 com.google.common.cache
[ИНФОРМАЦИЯ] 8478 com.google.common.collect
[ИНФОРМАЦИЯ] 50 com.google.common.eventbus
[ИНФОРМАЦИЯ] 385 com.google.common.hash
[ИНФОРМАЦИЯ] 597 com.google.common.io
[ИНФОРМАЦИЯ] 92 com.google.common.math
[ИНФОРМАЦИЯ] 134 com.google.common.net
[ИНФОРМАЦИЯ] 521 com.google.common.primitives
[ИНФОРМАЦИЯ] 404 com.google.common.reflect
[ИНФОРМАЦИЯ] 954 com.google.common.util.concurrent
[ИНФОРМАЦИЯ] 16 по умолчанию
[ИНФОРМАЦИЯ] 227 it. unimi.dsi.fastutil
[ИНФОРМАЦИЯ] 1370 ит.уними.dsi.fastutil.booleans
[ИНФОРМАЦИЯ] 17249 it.unimi.dsi.fastutil.bytes
[ИНФОРМАЦИЯ] 17249 it.unimi.dsi.fastutil.chars
[ИНФОРМАЦИЯ] 17279 it.unimi.dsi.fastutil.doubles
[ИНФОРМАЦИЯ] 17263 it.unimi.dsi.fastutil.floats
[ИНФОРМАЦИЯ] 17309 it.unimi.dsi.fastutil.ints
[ИНФОРМАЦИЯ] 628 it.unimi.dsi.fastutil.io
[ИНФОРМАЦИЯ] 17315 it.unimi.dsi.fastutil.longs
[ИНФОРМАЦИЯ] 26514 it.unimi.dsi.fastutil.objects
[ИНФОРМАЦИЯ] 17257 it.unimi.dsi.fastutil.shorts
[ИНФОРМАЦИЯ] 2 java.awt
[ИНФОРМАЦИЯ] 5 java.awt.datatransfer
[ИНФОРМАЦИЯ] 206 java.io
[ИНФОРМАЦИЯ] 346 java.язык
[ИНФОРМАЦИЯ] 10 java.lang.ref
[ИНФОРМАЦИЯ] 71 java.lang.reflect
[ИНФОРМАЦИЯ] 27 java.math
[ИНФОРМАЦИЯ] 26 java.net
[ИНФОРМАЦИЯ] 108 java.nio
[ИНФОРМАЦИЯ] 6 java.nio.channels
[ИНФОРМАЦИЯ] 2 java.nio.charset
[ИНФОРМАЦИЯ] 13 java.security
[ИНФОРМАЦИЯ] 2 java.text
[ИНФОРМАЦИЯ] 382 java.util
[ИНФОРМАЦИЯ] 128 java.util.concurrent
[ИНФОРМАЦИЯ] 33 java.util.concurrent.atomic
[ИНФОРМАЦИЯ] 40 java.util.concurrent.
unimi.dsi.fastutil
[ИНФОРМАЦИЯ] 1370 ит.уними.dsi.fastutil.booleans
[ИНФОРМАЦИЯ] 17249 it.unimi.dsi.fastutil.bytes
[ИНФОРМАЦИЯ] 17249 it.unimi.dsi.fastutil.chars
[ИНФОРМАЦИЯ] 17279 it.unimi.dsi.fastutil.doubles
[ИНФОРМАЦИЯ] 17263 it.unimi.dsi.fastutil.floats
[ИНФОРМАЦИЯ] 17309 it.unimi.dsi.fastutil.ints
[ИНФОРМАЦИЯ] 628 it.unimi.dsi.fastutil.io
[ИНФОРМАЦИЯ] 17315 it.unimi.dsi.fastutil.longs
[ИНФОРМАЦИЯ] 26514 it.unimi.dsi.fastutil.objects
[ИНФОРМАЦИЯ] 17257 it.unimi.dsi.fastutil.shorts
[ИНФОРМАЦИЯ] 2 java.awt
[ИНФОРМАЦИЯ] 5 java.awt.datatransfer
[ИНФОРМАЦИЯ] 206 java.io
[ИНФОРМАЦИЯ] 346 java.язык
[ИНФОРМАЦИЯ] 10 java.lang.ref
[ИНФОРМАЦИЯ] 71 java.lang.reflect
[ИНФОРМАЦИЯ] 27 java.math
[ИНФОРМАЦИЯ] 26 java.net
[ИНФОРМАЦИЯ] 108 java.nio
[ИНФОРМАЦИЯ] 6 java.nio.channels
[ИНФОРМАЦИЯ] 2 java.nio.charset
[ИНФОРМАЦИЯ] 13 java.security
[ИНФОРМАЦИЯ] 2 java.text
[ИНФОРМАЦИЯ] 382 java.util
[ИНФОРМАЦИЯ] 128 java.util.concurrent
[ИНФОРМАЦИЯ] 33 java.util.concurrent.atomic
[ИНФОРМАЦИЯ] 40 java.util.concurrent. locks
[ИНФОРМАЦИЯ] 6 java.util.jar
[ИНФОРМАЦИЯ] 6 java.util.logging
[ИНФОРМАЦИЯ] 10 java.util.regex
[ИНФОРМАЦИЯ] 20 java.util.zip
[ИНФОРМАЦИЯ] 26 javax.microedition.khronos.egl
[ИНФОРМАЦИЯ] 188 javax.microedition.khronos.opengles
[ИНФОРМАЦИЯ] 7 вс.misc
locks
[ИНФОРМАЦИЯ] 6 java.util.jar
[ИНФОРМАЦИЯ] 6 java.util.logging
[ИНФОРМАЦИЯ] 10 java.util.regex
[ИНФОРМАЦИЯ] 20 java.util.zip
[ИНФОРМАЦИЯ] 26 javax.microedition.khronos.egl
[ИНФОРМАЦИЯ] 188 javax.microedition.khronos.opengles
[ИНФОРМАЦИЯ] 7 вс.misc
Как видите, в классах fastutil огромное количество ссылок.
Есть два возможных решения этой проблемы:
- Используйте альтернативу проблемной зависимости (да).
- Используйте оптимизатор, например Proguard (общий для Android, только для Maven).
Внутрипроектный репозиторий Maven, вызывающий ошибки зависимости
Эта ошибка Maven полностью поставила меня в тупик, поэтому мне нужна помощь.У меня есть проект, в котором объявляется внутрипроектное репо:
<родитель>
ca.gc.agr.common
сухой док-родитель
../DryDockPOM/pom.xml
CognosReportService
<упаковка> банка
2. 0
CognosReportService
<репозитории>
<репозиторий>
в проекте
В репозитории проекта
file: // $ {project.basedir} / libs
<зависимости>
... вещи ...
<зависимость>
cognos-sdk
когнос-клиент
8.4.1
<зависимость>
cognos-sdk
когнос-ось
8.4.1
... вещи ...
 0
0 Есть еще один проект, который использует его так:
<родитель>
ca.gc.agr.common
общий веб-родитель
../DryDockWebPOM/pom.xml
CognosReportWeb
<упаковка> банка
2. 0
CognosReportWeb
<зависимости>
... вещи ....
<зависимость>
ca.gc.agr.common
CognosReportService
[1.0,)
... вещи ....
 0
0 Если я перейду к родительскому проекту и сделаю mvn install оттуда, все заработает. Однако, если я попытаюсь собрать CognosReportWeb самостоятельно, я получу следующее:
[ОШИБКА] Не удалось выполнить цель проекта CognosReportWeb: не удалось разрешить зависимости для проекта ca.gc.agr.common: CognosReportWeb: jar: 2.0:
Не удалось собрать зависимости для [ca.gc.agr.common: CognosReportService: jar: [1.0,) (компиляция),
ca.gc.agr.common: CommonWebJar: jar: [1.0,) (компилировать),
org.apache.tomcat: tomcat-servlet-api: jar: 7.0.16 (предоставляется),
javax.servlet.jsp: jsp-api: jar: 2.2 (предоставляется),
javax.el: el-api: jar: 2.2 (предоставляется),
org. glassfish.web: el-impl: jar: 2.2 (прилагается),
com.sun.faces: jsf-api: jar: 2.1.7 (компилировать),
com.sun.faces: jsf-impl: jar: 2.1.7 (компилировать),
org.apache.myfaces.tomahawk: tomahawk20: jar: 1.1.10 (компилировать),
org.richfaces: richfaces-bom: pom: 4.2.0.Final (компилировать),
org.richfaces.ui: richfaces-components-ui: jar: 4.2.0.Final (компиляция),
org.richfaces.core: richfaces-core-impl: jar: 4.2.0.Final (компиляция),
org.primefaces: primefaces: jar: 5.0 (компилировать),
org.springframework: spring-web: jar: 4.0.0.RELEASE (скомпилировать),
com.cybozu.labs: langdetect: jar: 1.1-20120112 (компилировать),
org.apache.myfaces.test: myfaces-test20: jar: 1.0.7 (тест),
org.aspectj: aspectjweaver: jar: 1.7.4 (компиляция),
org.springframework: контекст-пружины: jar: 4.0.0.RELEASE (компилировать),
org.springframework: аспекты-пружины: jar: 4.0.0.RELEASE (компиляция),
org.springframework: spring-tx: jar: 4.0.0.RELEASE (компилировать),
org.springframework: spring-test: jar: 4. 0.0.RELEASE (компилировать),
org.springframework: spring-orm: jar: 4.0.0.RELEASE (компилировать),
org.springframework: spring-oxm: jar: 4.0.0.RELEASE (компилировать),
org.springframework: spring-jdbc: jar: 4.0.0.RELEASE (компилировать),
org.hibernate: hibernate-core: jar: 4.1.7.Final (компилировать),
commons-io: commons-io: jar: 1.3.2 (компилировать),
Commons-lang: commons-lang: jar: 2.4 (компилировать),
commons-beanutils: commons-beanutils: jar: 1.8.3 (компиляция),
Commons-validator: commons-validator: jar: 1.3.1 (компилировать),
log4j: log4j: jar: 1.2.17 (компилировать),
org.slf4j: slf4j-log4j12: jar: 1.7.6 (компилировать),
junit: junit: jar: 4.11 (компилировать),
junit-addons: junit-addons: jar: 1.4 (компилировать),
com.googlecode.log4jdbc: log4jdbc: jar: 1.2 (компиляция),
org.reflections: отражения: jar: 0.9.9-RC1 (компилировать),
oro: oro: jar: 2.0.8 (компилировать),
dbunit: dbunit: jar: 2.1 (компилировать),
com.h3database: h3: jar: 1.3.175 (компилировать)]:
Не удалось прочитать дескриптор артефакта для организации. аспектj: аспектjweaver: jar: 1.8.0.M1:
Не удалось передать артефакт org.aspectj: aspectjweaver: pom: 1.8.0.M1 из / в проект (файл: // $ {project.basedir} / libs):
Путь к репозиторию / библиотеки не существует и не может быть создан. -> [Справка 1]
 glassfish.web: el-impl: jar: 2.2 (прилагается),
com.sun.faces: jsf-api: jar: 2.1.7 (компилировать),
com.sun.faces: jsf-impl: jar: 2.1.7 (компилировать),
org.apache.myfaces.tomahawk: tomahawk20: jar: 1.1.10 (компилировать),
org.richfaces: richfaces-bom: pom: 4.2.0.Final (компилировать),
org.richfaces.ui: richfaces-components-ui: jar: 4.2.0.Final (компиляция),
org.richfaces.core: richfaces-core-impl: jar: 4.2.0.Final (компиляция),
org.primefaces: primefaces: jar: 5.0 (компилировать),
org.springframework: spring-web: jar: 4.0.0.RELEASE (скомпилировать),
com.cybozu.labs: langdetect: jar: 1.1-20120112 (компилировать),
org.apache.myfaces.test: myfaces-test20: jar: 1.0.7 (тест),
org.aspectj: aspectjweaver: jar: 1.7.4 (компиляция),
org.springframework: контекст-пружины: jar: 4.0.0.RELEASE (компилировать),
org.springframework: аспекты-пружины: jar: 4.0.0.RELEASE (компиляция),
org.springframework: spring-tx: jar: 4.0.0.RELEASE (компилировать),
org.springframework: spring-test: jar: 4.
glassfish.web: el-impl: jar: 2.2 (прилагается),
com.sun.faces: jsf-api: jar: 2.1.7 (компилировать),
com.sun.faces: jsf-impl: jar: 2.1.7 (компилировать),
org.apache.myfaces.tomahawk: tomahawk20: jar: 1.1.10 (компилировать),
org.richfaces: richfaces-bom: pom: 4.2.0.Final (компилировать),
org.richfaces.ui: richfaces-components-ui: jar: 4.2.0.Final (компиляция),
org.richfaces.core: richfaces-core-impl: jar: 4.2.0.Final (компиляция),
org.primefaces: primefaces: jar: 5.0 (компилировать),
org.springframework: spring-web: jar: 4.0.0.RELEASE (скомпилировать),
com.cybozu.labs: langdetect: jar: 1.1-20120112 (компилировать),
org.apache.myfaces.test: myfaces-test20: jar: 1.0.7 (тест),
org.aspectj: aspectjweaver: jar: 1.7.4 (компиляция),
org.springframework: контекст-пружины: jar: 4.0.0.RELEASE (компилировать),
org.springframework: аспекты-пружины: jar: 4.0.0.RELEASE (компиляция),
org.springframework: spring-tx: jar: 4.0.0.RELEASE (компилировать),
org.springframework: spring-test: jar: 4. 0.0.RELEASE (компилировать),
org.springframework: spring-orm: jar: 4.0.0.RELEASE (компилировать),
org.springframework: spring-oxm: jar: 4.0.0.RELEASE (компилировать),
org.springframework: spring-jdbc: jar: 4.0.0.RELEASE (компилировать),
org.hibernate: hibernate-core: jar: 4.1.7.Final (компилировать),
commons-io: commons-io: jar: 1.3.2 (компилировать),
Commons-lang: commons-lang: jar: 2.4 (компилировать),
commons-beanutils: commons-beanutils: jar: 1.8.3 (компиляция),
Commons-validator: commons-validator: jar: 1.3.1 (компилировать),
log4j: log4j: jar: 1.2.17 (компилировать),
org.slf4j: slf4j-log4j12: jar: 1.7.6 (компилировать),
junit: junit: jar: 4.11 (компилировать),
junit-addons: junit-addons: jar: 1.4 (компилировать),
com.googlecode.log4jdbc: log4jdbc: jar: 1.2 (компиляция),
org.reflections: отражения: jar: 0.9.9-RC1 (компилировать),
oro: oro: jar: 2.0.8 (компилировать),
dbunit: dbunit: jar: 2.1 (компилировать),
com.h3database: h3: jar: 1.3.175 (компилировать)]:
Не удалось прочитать дескриптор артефакта для организации.
0.0.RELEASE (компилировать),
org.springframework: spring-orm: jar: 4.0.0.RELEASE (компилировать),
org.springframework: spring-oxm: jar: 4.0.0.RELEASE (компилировать),
org.springframework: spring-jdbc: jar: 4.0.0.RELEASE (компилировать),
org.hibernate: hibernate-core: jar: 4.1.7.Final (компилировать),
commons-io: commons-io: jar: 1.3.2 (компилировать),
Commons-lang: commons-lang: jar: 2.4 (компилировать),
commons-beanutils: commons-beanutils: jar: 1.8.3 (компиляция),
Commons-validator: commons-validator: jar: 1.3.1 (компилировать),
log4j: log4j: jar: 1.2.17 (компилировать),
org.slf4j: slf4j-log4j12: jar: 1.7.6 (компилировать),
junit: junit: jar: 4.11 (компилировать),
junit-addons: junit-addons: jar: 1.4 (компилировать),
com.googlecode.log4jdbc: log4jdbc: jar: 1.2 (компиляция),
org.reflections: отражения: jar: 0.9.9-RC1 (компилировать),
oro: oro: jar: 2.0.8 (компилировать),
dbunit: dbunit: jar: 2.1 (компилировать),
com.h3database: h3: jar: 1.3.175 (компилировать)]:
Не удалось прочитать дескриптор артефакта для организации. аспектj: аспектjweaver: jar: 1.8.0.M1:
Не удалось передать артефакт org.aspectj: aspectjweaver: pom: 1.8.0.M1 из / в проект (файл: // $ {project.basedir} / libs):
Путь к репозиторию / библиотеки не существует и не может быть создан. -> [Справка 1]
аспектj: аспектjweaver: jar: 1.8.0.M1:
Не удалось передать артефакт org.aspectj: aspectjweaver: pom: 1.8.0.M1 из / в проект (файл: // $ {project.basedir} / libs):
Путь к репозиторию / библиотеки не существует и не может быть создан. -> [Справка 1]
На первый взгляд кажется, что есть проблема с org.aspectj: Aspectjweaver: pom: 1.8.0.M1 , но я, если я закомментирую собственное внутрипроектное репо в CognosReportService (и повторно -install), вдруг CognosReportWeb начинает работать.
Я действительно в тупике. Я делаю что-то неправильно?
Будем признательны за любые идеи!
ДНК-зависимая архитектура хроматина и образование ядерных узлов
Деккер, Дж. И Херд, Э. Структурное и функциональное разнообразие топологически ассоциированных доменов. FEBS Lett. 589 , 2877–2884 (2015).
CAS PubMed PubMed Central Статья Google ученый
Секстон Т. и Кавалли Г. Роль хромосомных доменов в формировании функционального генома. Cell. 160 , 1049–59 (2015).
CAS PubMed Статья PubMed Central Google ученый
Ю. М. и Рен Б. Трехмерная организация геномов млекопитающих. Annu Rev Cell Dev Biol. 33 , 265–289 (2017).
CAS PubMed PubMed Central Статья Google ученый
Роули, М. Дж. И Корсес, В. Г. Организационные принципы трехмерной архитектуры генома. Nat Rev Genet. 19 , 789–800 (2018).
Nat Rev Genet. 19 , 789–800 (2018).
CAS PubMed Статья PubMed Central Google ученый
van Steensel, B. & Furlong, E. E. M. Роль транскрипции в формировании пространственной организации генома. Nat Rev Mol Cell Biol 20 , 327–337 (2019).
PubMed Статья CAS PubMed Central Google ученый
Диксон, Дж. Р. и др. . Топологические домены в геномах млекопитающих, идентифицированные путем анализа взаимодействий хроматина. Природа. 485 , 376–380 (2012).
CAS PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google ученый
Нора, Э. П. и др. . Пространственное разделение регуляторного ландшафта центра X-инактивации. Природа. 485 , 381–385 (2012).
CAS PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google ученый
Дауэн, Дж. М. и др. . Контроль генов клеточной идентичности происходит в изолированных окрестностях хромосом млекопитающих. Cell. 159 , 374–387 (2014).
CAS PubMed PubMed Central Статья Google ученый
Phillips-Cremins, J.Е. и др. . Подклассы архитектурных белков формируют трехмерную организацию геномов во время фиксации клонов. Cell. 153 , 1281–95 (2013).
CAS PubMed PubMed Central Статья Google ученый
 org/ScholarlyArticle»> 10.
org/ScholarlyArticle»> 10.Рао, С. С. П. и др. . Трехмерная карта генома человека с разрешением в килобазы раскрывает принципы образования петель хроматина. Cell. 159 , 1665–1680 (2014).
CAS PubMed PubMed Central Статья Google ученый
Джи, Х. и др. . Трехмерный регуляторный ландшафт хромосом плюрипотентных клеток человека. Cell. 18 , 262–275 (2016).
CAS Google ученый
Guelen, L. et al. . Конститутивные взаимодействия ядерной ламины и генома высококонсервативны и связаны с A / T-богатой последовательностью. Genome Res. 2 , 270–80 (2013).
Google ученый
 org/ScholarlyArticle»> 13.
org/ScholarlyArticle»> 13.Jabbari, K. & Bernardi, G. Каркас изохор лежит в основе архитектуры хроматина. PLoS One. 12 , e0168023 (2017).
PubMed PubMed Central Статья CAS Google ученый
Нотон, К. и др. . Транскрипция формирует и ремоделирует суперспиральные домены, разворачивая крупномасштабные структуры хроматина. Nat Struct Mol Biol. 20 , 387–95 (2013).
CAS PubMed PubMed Central Статья Google ученый
Корлесс, С. и Гилберт, Н. Эффекты суперспирализации ДНК на архитектуру хроматина. Biophys Rev. 8 , 245–258 (2016).
CAS PubMed PubMed Central Статья Google ученый
 org/ScholarlyArticle»> 16.
org/ScholarlyArticle»> 16.Peric-Hupkes, D. et al. . Mol Cell. 38 , 603–13 (2010).
CAS PubMed PubMed Central Статья Google ученый
ван Стинзель, Б. и Белмонт, А. С. Домены, связанные с ламинами: связи с архитектурой хромосомы, гетерохроматином и репрессией генов. Cell. 169 , 780–791 (2017).
PubMed PubMed Central Статья CAS Google ученый
Чандра, Т. и др. . Глобальная реорганизация ядерного ландшафта в стареющих клетках. Cell Rep. 10 , 471–83 (2015).
CAS PubMed PubMed Central Статья Google ученый
 org/ScholarlyArticle»> 19.
org/ScholarlyArticle»> 19.Quinodoz, S.A. et al. . Межхромосомные центры высшего порядка формируют трехмерную организацию генома в ядре. Cell. 174 , 744–757 (2018).
CAS PubMed PubMed Central Статья Google ученый
Лонг, Х. К., Прескотт, С. Л. и Высока, Дж. Постоянно меняющиеся ландшафты: усилители транскрипции в развитии и эволюции. Cell. 167 , 1170–1187 (2016).
CAS PubMed PubMed Central Статья Google ученый
Галупа, Р. и Херд, Э. Топологически связывающие домены в архитектуре хромосом и регуляторных ландшафтах генов во время развития, болезней и эволюции. Колд Спринг Харб Symp Quant Biol. 82 , 267–278 (2017).
PubMed Статья PubMed Central Google ученый
Банани, С. Ф., Ли, Х. О., Хайман, А. А. и Розен, М. К. Биомолекулярные конденсаты: организаторы клеточной биохимии. Nat Rev Mol Cell Biol. 18 , 285–298 (2017).
CAS PubMed Статья PubMed Central Google ученый
Fenouil, R. et al. . CpG-островки и содержание GC диктуют истощение нуклеосом независимым от транскрипции образом на промоторах млекопитающих. Genome Res. 22 , 2399–2408 (2012).
CAS PubMed PubMed Central Статья Google ученый
 org/ScholarlyArticle»> 24.
org/ScholarlyArticle»> 24.Struhl, K. & Segal, E. Детерминанты расположения нуклеосом. Nature Struct и Mol Biol. 20 , 267–273 (2013).
CAS Статья Google ученый
Дриллон, Г., Аудит, Б., Аргул, Ф. и Арнеодо, А. Доказательства отбора доступного массива нуклеосом у человека. BMC Genomics. 17 , 526 (2016).
PubMed PubMed Central Статья CAS Google ученый
Калхор, Р., Тьонг, Х., Джаятилака, Н., Альбер, Ф. и Чен, Л.Архитектура генома, выявленная с помощью фиксации конформации привязанной хромосомы и популяционного моделирования. Nat Biotechnol. 30 , 90–98 (2011).
PubMed PubMed Central Статья CAS Google ученый
 org/ScholarlyArticle»> 27.
org/ScholarlyArticle»> 27.Tjong, H. et al. . Популяционный анализ трехмерной структуры генома выявляет движущие силы в пространственной организации генома. Proc Natl Acad Sci USA 113 , E1663–1672 (2016).
CAS PubMed Статья PubMed Central Google ученый
Клэй, О., Карелс, Н., Дуади, К., Макайя, Г. и Бернарди, Г. Композиционная неоднородность внутри и между изохорами в геномах млекопитающих. I. CsCl и анализ последовательностей. Gene. 276 , 15–24 (2001).
CAS PubMed Статья PubMed Central Google ученый
Zufferey, M., Tavernari, D., Oricchio, E. & Ciriello, G. Сравнение вычислительных методов для идентификации топологически связанных областей. Genome Biol. 19 , 217 (2018).
Genome Biol. 19 , 217 (2018).
CAS PubMed PubMed Central Статья Google ученый
Мэтьюз, Б. Дж. И Ваксман, Д. Дж. Вычислительное предсказание основанных на CTCF / когезине петель внутри-TAD, которые изолируют контакты хроматина и экспрессию генов в печени мыши. Элиф. 7 , e34077 (2018).
PubMed PubMed Central Статья Google ученый
Caudron-Herger, M. et al. . РНК, содержащие элементы Alu, поддерживают структуру и функцию ядрышка. EMBO J. 34 , 2758–2574 (2015).
CAS PubMed PubMed Central Статья Google ученый
 org/ScholarlyArticle»> 32.
org/ScholarlyArticle»> 32.Шин, Ю. и др. . Жидкие ядерные конденсаты механически чувствуют и реструктурируют геном. Cell. 175 , 1481–1491.e13 (2018).
CAS PubMed Статья PubMed Central Google ученый
Saccone, S., Federico, C. & Bernardi, G. Локализация наиболее богатых генами и самых бедных генами изохор в интерфазных ядрах млекопитающих и птиц. Gene. 300 , 169–78 (2002).
CAS PubMed Статья PubMed Central Google ученый
Cremer, T. et al. . Четырехмерный нуклеом: свидетельство динамического ядерного ландшафта, основанного на совместных активных и неактивных ядерных компартментах. FEBS Lett. 589 , 2931–43 (2015).
CAS PubMed Статья PubMed Central Google ученый
Yaffe, E. & Tanay, A. Вероятностное моделирование карт контактов Hi-C устраняет систематические искажения для характеристики глобальной хромосомной архитектуры. Nat Genet. 43 , 1059–1065 (2011).
CAS PubMed Статья PubMed Central Google ученый
Olivares-Chauvet, P. et al. . Захват попарных и многосторонних хромосомных конформаций с помощью хромосомных блужданий. Природа. 540 , 296–300 (2016).
CAS PubMed Статья ОБЪЯВЛЕНИЯ PubMed Central Google ученый
 org/ScholarlyArticle»> 37.
org/ScholarlyArticle»> 37.Виноградов А.Е. Спираль ДНК: важность ГХ-обогащения. Nucleic Acids Res. 31 , 1838–44 (2003).
CAS PubMed PubMed Central Статья Google ученый
Джаббари К. и Бернарди Г.Метилирование цитозина и частоты CpG, TpG (CpA) и TpA. Gene. 333 , 143–149 (2004).
CAS PubMed Статья PubMed Central Google ученый
Li, W. & Miramontes, P. Крупномасштабные колебания структурно-зависимых характеристик последовательности ДНК в хромосоме 21 человека. Phys Rev E Stat Nonlin Soft Matter Phys. 74 , 021912 (2006).
PubMed Статья ОБЪЯВЛЕНИЯ CAS PubMed Central Google ученый
 org/ScholarlyArticle»> 40.
org/ScholarlyArticle»> 40.Пекхэм, Х. Э. и др. . Сигналы позиционирования нуклеосом в геномной ДНК. Genome Res. 17 , 1170–1177 (2007).
CAS PubMed PubMed Central Статья Google ученый
Локк, Г., Толкунов, Д., Моктадери, З., Струль, К. и Морозов, А. В. Высокопроизводительное секвенирование раскрывает простую модель энергетики нуклеосом. Proc Natl Acad Sci USA 107 , 20998–1003 (2010).
CAS PubMed Статья ОБЪЯВЛЕНИЯ Google ученый
Чуанг, Х. М., Райфенбергер, Дж. Г., Цао, Х. и Дорфман, К. Д. Последовательно-зависимая стойкость. Длина длинной ДНК. Phys Rev Lett. 119 , 227802 (2017).
PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google ученый
 org/ScholarlyArticle»> 43.
org/ScholarlyArticle»> 43.Riggs, A. D. Метилирование ДНК и поздняя репликация, вероятно, способствуют клеточной памяти, а наматывание ДНК типа I может способствовать укладке хромосом и функции энхансера. Philos Trans R Soc Lond Biol Sci. 326 , 285–297 (1990).
CAS Статья ОБЪЯВЛЕНИЯ Google ученый
Нэсмит, К. Распространение генома: соединение, разрешение и разделение сестринских хроматид во время митоза и мейоза. Annu Rev Genet. 35 , 673–745 (2001).
CAS PubMed Статья PubMed Central Google ученый
Алипур, Э. и Марко, Дж. Ф. Самоорганизация доменных структур ферментами, выдавливающими петли ДНК. Nucleic Acids Res. 40 , 11202–11212 (2012).
40 , 11202–11212 (2012).
CAS PubMed PubMed Central Статья Google ученый
Голобородько А., Марко Дж. Ф. и Мирный Л. А. Уплотнение хромосом с помощью активной экструзии петель. Biophys J. 110 , 2162–2168 (2016).
CAS PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google ученый
Баррингтон, К., Финн, Р. и Хаджур, С. Биология Кохесина соответствует модели вытеснения петли. Chromosome Res. 25 , 51–60 (2017).
CAS PubMed PubMed Central Статья Google ученый

Terakawa, T. et al. . Конденсирующий комплекс — это механохимический двигатель, который перемещается вдоль ДНК. Наука. 358 , 672–676 (2017).
CAS PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google ученый
Нэсмит, К. Как ДНК вплетены в хромосомы? Наука. 358 , 589–590 (2017).
CAS PubMed Статья ОБЪЯВЛЕНИЯ PubMed Central Google ученый
Vian, L. et al. . Энергетика и физиологическое влияние экструзии когезина. Cell. 175 , 292–294 (2018).
CAS PubMed PubMed Central Статья Google ученый
Нишияма, Т. Сплоченность и когезин-зависимая организация хроматина. Curr Opin Cell. Биол. 58 , 8–14 (2018).
PubMed Статья CAS PubMed Central Google ученый
Ea, V. et al. . Четкие принципы физики полимеров управляют динамикой хроматина в топологических доменах мышей и дрозофилы. BMC Genomics. 16 , 607 (2015).
PubMed PubMed Central Статья CAS Google ученый
Торрес, К. М. и др. . Линкерный гистон h2.0 создает эпигенетическую и функциональную внутриопухолевую гетерогенность. Наука. 353 , 6307 (2016).
Артикул CAS Google ученый
Клекнер, Н. Формирование хиазмы: взаимодействие хроматина / оси и роль (и) синаптонемного комплекса. Хромосома 115 , 175–194 (2006).
PubMed Статья PubMed Central Google ученый
Джаббари, К., Виртц, Дж., Раушер, М. и Вие, Т. Общий геномный код для архитектуры хроматина и ландшафта рекомбинации. PLoS One. 14 , e0213278 (2019).
CAS PubMed PubMed Central Статья Google ученый
Диксон, Дж. Р., Горкин, Д. У. и Рен, Б. Домены хроматина: единица хромосомной организации. Mol Cell. 62 , 668–80 (2016).
CAS PubMed PubMed Central Статья Google ученый
Bernardi, G. Формирование доменов хроматина включает первичный этап, основанный на трехмерной структуре ДНК. Научная корреспонденция 8 , 17821 (2018).
PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ CAS Google ученый
Чжан, Х., Бранчяморе, С., Гогошин, Г., Родин, А. С. и Риггс, А. Д. Анализ трехмерных внутрихромосомных взаимодействий с высоким разрешением с помощью моделирования байесовской сети. Proc Natl Acad Sci USA 114 , E10359 – E10368 (2017).
CAS PubMed Статья PubMed Central Google ученый
Zuin, J. et al. . Cohesin и CTCF по-разному влияют на архитектуру хроматина и экспрессию генов в клетках человека. Proc Natl Acad Sci USA 111 , 996–1001 (2014).
CAS PubMed Статья ОБЪЯВЛЕНИЯ PubMed Central Google ученый
Schwarzer, W. et al. . При удалении когезина выявлены два независимых способа организации хроматина. Природа. 551 , 51–56 (2017).
PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google ученый
Рао, С. С. П. и др. .Потеря Cohesin исключает все петлевые домены. Cell. 171 , 305–320.e24 (2017).
CAS PubMed PubMed Central Статья Google ученый
Нора, Э. П. и др. . Направленная деградация CTCF отделяет локальную изоляцию хромосомных доменов от компартментализации генома. Cell. 169 , 930–944.e22 (2017).
CAS PubMed PubMed Central Статья Google ученый
Уускюла-Рейманд, Л. и др. . Топоизомераза II бета взаимодействует с когезином и CTCF на границах топологических доменов. Genome Biol. 17 , 182 (2016).
PubMed PubMed Central Google ученый
Рако, Д., Бенедетти, Ф., Дориер, Дж. И Стасиак, А. Являются ли TAD суперспиралями? Nucleic Acids Res. 47 , 521–532 (2019).
CAS PubMed Статья PubMed Central Google ученый
Гот, Х. Дж. и др. . Пространственное сворачивание хромосом и активная транскрипция управляют хрупкостью ДНК и образованием онкогенных транслокаций MLL. Mol Cell. 75 , 267–283 (2019).
CAS PubMed Статья PubMed Central Google ученый
Хайман А. А., Вебер К. А. и Юлихер Ф. Разделение жидких и жидких фаз в биологии. Annu Rev Cell Dev Biol. 30 , 39–58 (2014).
CAS PubMed Статья PubMed Central Google ученый
Cho, W. K. et al. . Кластеры медиатора и РНК-полимеразы II объединяются в зависимые от транскрипции конденсаты. Наука. 361 , 412–415 (2018).
CAS PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google ученый
Сабари, Б.Р. и др. . Конденсация коактиватора на суперэнхансерах связывает разделение фаз и контроль генов. Наука. 361 , 6400 (2018).
Артикул CAS Google ученый
Németh, A. et al. . Первоначальная геномика ядрышка человека. PLoS Genet. 6 , e1000889 (2010).
PubMed PubMed Central Статья CAS Google ученый
Ли, П. и др. . Фазовые переходы в сборке поливалентных сигнальных белков. Природа. 483 , 336–40 (2012).
CAS PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google ученый
МакИнтош, Д. Б., Дагган, Г., Гуил, К. и Салех, О. А. Последовательно-зависимая эластичность и электростатика одноцепочечной ДНК: признаки укладки оснований. Biophys J. 106 , 659–666 (2014).
CAS PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google ученый
Шакья, А. и Кинг, Дж. Т. Сборка разделенных фаз жидких капель ДНК, зависящая от локальной гибкости. Biophys J. 115 , 1840–1847 (2018).
CAS PubMed Статья ОБЪЯВЛЕНИЯ PubMed Central Google ученый
Peters, J. P.3rd & Maher, L.J. Кривизна и гибкость ДНК in vitro и in vivo . Q Rev Biophys. 43 , 23–63 (2010).
CAS PubMed PubMed Central Статья Google ученый
Ricci, M.A., Manzo, C., García-Parajo, M. F., Lakadamyali, M. & Cosma, M. P. Хроматиновые волокна образованы гетерогенными группами нуклеосом in vivo . Cell. 160 , 1145–1158 (2015).
CAS PubMed Статья PubMed Central Google ученый
Boija, A. et al. . Факторы транскрипции активируют гены за счет способности их активационных доменов к разделению фаз. Cell. 175 , 1842–1855 (2018).
CAS PubMed Статья PubMed Central Google ученый
Хниш, Д., Шринивас, К., Янг, Р. А., Чакраборти, А. К. и Шарп, П. А. Модель разделения фаз для контроля транскрипции. Cell. 23 , 13–23 (2017).
Артикул CAS Google ученый
Циркель А. и Папантонис А. Транскрипция как сила, разделяющая геном эукариот. Biol Chem. 395 , 1301–5 (2014).
CAS PubMed Статья PubMed Central Google ученый
Трифонов Е. Н. Множественные коды нуклеотидных последовательностей. Bull Math Biol. 5 , 417–32 (1989).
MATH Статья Google ученый
Граур Д. Верхний предел функциональной части генома человека. Genome Biol Evol. 9 , 1880–1885 (2017).
PubMed PubMed Central Статья Google ученый
Дулиттл, В. Ф. и Брюнет, Т. Д. П. О причинных ролях и избранных эффектах: наш геном в основном мусор. BMC Biol. 15 , 116 (2017).
PubMed PubMed Central Статья CAS Google ученый
Цукеркандл, Е. Политическая ДНК: функциональная плотность и функциональная совместимость в геномах. J Mol Evol. 24 , 12–27 (1986).
CAS PubMed Статья ОБЪЯВЛЕНИЯ PubMed Central Google ученый
Поуп, Б. Д. и др. . Топологически связанные домены представляют собой стабильные единицы регуляции времени репликации. Природа. 515 , 402–405 (2014).
CAS PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google ученый
Costantini, M., Clay, O., Auletta, F. & Bernardi, G. Карта изохор хромосом человека. Genome Res. 16 , 536–541 (2006).
CAS PubMed PubMed Central Статья Google ученый
Ле С., Джосс Дж. И Хассон Ф. FactoMineR: Пакет R для многомерного анализа. Журнал статистического программного обеспечения 25 , 1–18 (2008).
Артикул Google ученый
R Основная команда. R: Язык и среда для статистических вычислений. R Фонд статистических вычислений, Вена, Австрия. https://www.R-project.org/.
Кассамбара, А. и Мундт, Ф. Factoextra: извлечение и визуализация результатов многомерного анализа данных.Пакет R версии 1.0.5 (2017).
Харроу, Дж. и др. . GENCODE: справочная аннотация генома человека для проекта ENCODE. Genome Res. 22 , 1760–1774 (2012).
CAS PubMed PubMed Central Статья Google ученый
Khan, A. & Zhang, X. dbSUPER: база данных супер-энхансеров в геноме мыши и человека. Исследования нуклеиновых кислот. 44 , 164–171 (2016).
Артикул CAS Google ученый
Ziebarth, J., Bhattacharya, A. & Cui, Y. CTCFBSDB 2.0: база данных для сайтов связывания CTCF и организации генома. Исследования нуклеиновых кислот. 41 , 188–194 (2013).
Артикул CAS Google ученый
Куинлан, А. Р. и Холл, И. М. BEDTools: гибкий набор утилит для сравнения геномных характеристик. Биоинформатика. 26 , 841–842 (2010).
CAS PubMed PubMed Central Статья Google ученый
Крючкова-Мостаччи, Н. и Робинсон-Рехави, М. Контрольный показатель показателей тканевой специфичности экспрессии генов. Краткое. Биоинформ. 18 , 205–214 (2017).
CAS PubMed PubMed Central Google ученый
Смещение мутаций и инвазии антимутатора формы содержимого GC
Сниговски, П. Д., Герриш, П. Дж., Джонсон, Т. и Шейвер, А. Эволюция частоты мутаций: разделение причин и следствий. Bioessays 22 , 1057–1066 (2000).
CAS Статья Google ученый
Стертевант А. Х. Очерки эволюции. I. О влиянии отбора на частоту мутаций. Q. Rev. Biol. 12 , 464–467 (1937).
Артикул Google ученый
Кимура, М. Оптимальная частота мутаций и степень доминирования, определяемая по принципу минимальной генетической нагрузки. Дж. Жене . 57 , 21–34 (1960).
Артикул Google ученый
Ли, Э. Г. Эволюция скорости мутаций. Genetics 73 (Дополнение 73), 1–18 (1973).
ADS MathSciNet Google ученый
Эшел, И. Клон-отбор и оптимальные скорости мутации. J. Appl. Вероятно. 10 , 728–738 (1973).
MathSciNet Статья Google ученый
Либерман У. и Фельдман М. В. Модификаторы скорости мутаций: общий принцип редукции. Теор. Popul. Биол. 30 , 125–142 (1986).
MathSciNet CAS Статья Google ученый
Леклерк, Дж. Э., Ли, Б., Пейн, В. Л. и Себула, Т. А. Высокая частота мутаций среди патогенов Escherichia coli и Salmonella. Наука 274 , 1208–1211 (1996).
ADS CAS Статья Google ученый
Sniegowski, P. D., Gerrish, P. J. и Lenski, R. E. Эволюция высоких скоростей мутаций в экспериментальных популяциях E. coli. Nature 387 , 703–705 (1997).
ADS CAS Статья Google ученый
Хили К. Р., Хименес Ортигоса К., Шор Э. и Перлин Д. С. Генетические факторы множественной лекарственной устойчивости Candida glabrata. Фронт. Микробиол . 7 , 1995 (2016).
Voordeckers, K. et al. Адаптация к высокому этанолу показывает сложные эволюционные пути. PLOS Genet. 11 , e1005635 (2015).
Артикул Google ученый
Лоеб, Л. А. Рак человека выражает мутаторные фенотипы: происхождение, последствия и направленность. Nat. Rev. Cancer 11 , 450–457 (2011).
CAS Статья Google ученый
Гуд Б. Х. и Десаи М. М. Эволюция скорости мутаций в быстро адаптирующихся бесполых популяциях. Генетика 204 , 1249–1266 (2016).
Артикул Google ученый
Tenaillon, O., Toupance, B., Nagard, H.L., Taddei, F. & Godelle, B. Мутаторы, размер популяции, адаптивный ландшафт и адаптация бесполых популяций бактерий. Генетика 152 , 485–493 (1999).
CAS PubMed PubMed Central Google ученый
Тенайон О., Нагард Х. Л., Годелль Б. и Таддеи Ф. Мутаторы и пол у бактерий: конфликт между адаптивными стратегиями. PNAS 97 , 10465–10470 (2000).
ADS CAS Статья Google ученый
Raynes, Y., Wylie, C. S., Sniegowski, P. D. & Weinreich, D. M. Признак отбора по модификаторам скорости мутаций зависит от размера популяции. PNAS 115 , 3422–3427 (2018).
CAS Статья Google ученый
Дрейк, Дж.W. Общие антимутаторы маловероятны. J. Mol. Биол. 229 , 8–13 (1993).
CAS Статья Google ученый
Wielgoss, S. et al. Динамика скорости мутаций в бактериальной популяции отражает противоречие между адаптацией и генетической нагрузкой. Proc. Natl Acad. Sci. США 110 , 222–227 (2013).
ADS CAS Статья Google ученый
Brown, E. W., LeClerc, J. E., Li, B., Payne, W. L. и Cebula, T. A. Филогенетические доказательства горизонтального переноса аллелей mutS среди встречающихся в природе штаммов Escherichia coli. J. Bacteriol. 183 , 1631–1644 (2001).
CAS Статья Google ученый
Denamur, E. et al. Эволюционные последствия частого горизонтального переноса генов репарации несовпадений. Cell 103 , 711–721 (2000).
CAS Статья Google ученый
Елена, С.Ф., Уиттам, Т.С., Винкворт, К.Л., Райли, М.А. и Ленски, Р.Е. Геномная дивергенция штаммов Escherichia coli: доказательства горизонтального переноса и вариабельности частоты мутаций. Внутр. Microbiol. 8 , 271–278 (2005).
PubMed Google ученый
Couce, A., Alonso-Rodriguez, N., Костас, К., Оливер, А., Бласкес, Дж. Внутрипопуляционная изменчивость распространенности мутаторов среди изолятов инфекции мочевыводящих путей Escherichia coli. Clin. Microbiol. Заразить. 22 , 566.e1–7 (2016).
CAS Статья Google ученый
Макдональд, М. Дж., Се, Й.-Й., Ю, Й.-Х., Чанг, С.-Л. И Лей, Ж.-Й. Эволюция низких скоростей мутаций в экспериментальных популяциях мутаторов Saccharomyces cerevisiae. Curr. Биол. 22 , 1235–1240 (2012).
CAS Статья Google ученый
Сингх Т., Хюн М. и Сниговски П. Эволюция скорости мутаций в гипермутируемых популяциях Escherichia coli, размножающихся при очень небольшом эффективном размере популяции. Biol. Lett. 13 , 20160849 (2017).
Трёбнер В. и Пехоцки Р. Селекция против гипермутабильности у Escherichia coli в ходе долгосрочной эволюции. Molec Gen. Genet 198 , 177–178 (1984).
Артикул Google ученый
Turrientes, M.-C. и другие. Варианты с нормальной скоростью мутации возникают в популяции мутантной (Mut S) Escherichia coli. PLoS ONE 8 , e72963 (2013).
ADS CAS Статья Google ученый
Джейн, К. и Джеймс, А. Вероятность фиксации немутатора в большой популяции бесполых мутаторов. J. Theor. Биол. 433 , 85–93 (2017).
MathSciNet Статья Google ученый
Джеймс, А. и Джейн, К. Вероятность фиксации редкого немутатора и эволюция скорости мутаций. Ecol. Evol. 6 , 755–764 (2016).
Артикул Google ученый
Миллер, Дж. Х. Спонтанные мутаторы у бактерий: понимание путей мутагенеза и восстановления. Annu. Rev. Microbiol. 50 , 625–643 (1996).
CAS Статья Google ученый
Майклс, М. Л. и Миллер, Дж. Х. Система GO защищает организмы от мутагенного эффекта спонтанного поражения 8-гидроксигуанином (7,8-дигидро-8-оксогуанин). J. Bacteriol. 174 , 6321–6325 (1992).
CAS Статья Google ученый
Коус, А., Гуэльфо, Дж. Р. и Бласкес, Дж. Мутационный спектр стимулирует рост бактерий-мутаторов. PLoS Genet. 9 , e1003167 (2013).
CAS Статья Google ученый
Холдейн, Дж. Б. С. Эффект изменения приспособленности. Am. Nat. 71 , 337–349 (1937).
Артикул Google ученый
Мюллер, Х. Дж. Наш груз мутаций. Am. J. Hum. Genet 2 , 111–176 (1950).
CAS PubMed PubMed Central Google ученый
Кроу, Дж. Ф. Генетические нагрузки и стоимость естественного отбора. В «Математические темы в популяционной генетике» (редактор Кодзима, К.) 128–177 (Springer, Berlin Heidelberg, 1970).
Google ученый
Ямпольский, Л. Ю., Штольцфус, А. Обмениваемость аминокислот в белках. Генетика 170 , 1459–1472 (2005).
CAS Статья Google ученый
Гуд, Б. Х., Макдональд, М. Дж., Баррик, Дж. Э., Ленски, Р. Э. и Десаи, М. М. Динамика молекулярной эволюции на протяжении 60 000 поколений. Nature 551 , 45–50 (2017).
ADS Статья Google ученый
Фостер, П. Л. Методы определения частоты спонтанных мутаций. Meth. Энзим. 409 , 195–213 (2006).
CAS Статья Google ученый
Оливер А. и Мена А. Бактериальная гипермутация при муковисцидозе, а не только при устойчивости к антибиотикам. Clin. Microbiol. Заразить. 16 , 798–808 (2010).
CAS Статья Google ученый
Jolivet-Gougeon, A. et al. Бактериальная гипермутация: клинические последствия. J. Med. Microbiol. 60 , 563–573 (2011).
CAS Статья Google ученый
Notley-McRobb, L., Seeto, S. & Ferenci, T. Обогащение и устранение мутаторов mutY в популяциях Escherichia coli. Генетика 162 , 1055–1062 (2002).
CAS PubMed PubMed Central Google ученый
Ragheb, M. N. et al. Подавление развития устойчивости к антибиотикам. Мол. Ячейка 73 , 157–165.e5 (2019).
CAS Статья Google ученый
Эйр-Уокер, А. и Кейтли, П. Д. Распределение эффектов приспособляемости новых мутаций. Nat. Преподобный Жене. 8 , 610–618 (2007).
CAS Статья Google ученый
Елена, С. Ф. и Ленски, Р. Е. Тест синергетических взаимодействий между вредными мутациями у бактерий. Nature 390 , 395–398 (1997).
ADS CAS Статья Google ученый
Кибота Т. и Линч М. Оценка частоты геномных мутаций, вредных для общей пригодности E. coli. Nature 381 , 694–696 (1996).
ADS CAS Статья Google ученый
Trindade, S., Perfeito, L. & Gordo, I. Частота и эффекты спонтанных мутаций, которые влияют на приспособленность мутатора Escherichia coli. Philos Trans. R. Soc. Лондон. B Biol. Sci. 365 , 1177–1186 (2010).
Артикул Google ученый
Линд, П. А. и Андерссон, Д. И. Полногеномные мутационные искажения у бактерий. Proc. Natl Acad. Sci. США 105 , 17878–17883 (2008).
ADS CAS Статья Google ученый
Heilbron, K., Toll-Riera, M., Kojadinovic, M. & MacLean, R.C. Пригодность сильно зависит от редких мутаций большого эффекта в эксперименте по накоплению микробных мутаций. Генетика 197 , 981–990 (2014).
CAS Статья Google ученый
Диллон, М. и Купер, В. С. Эффекты приспособляемости спонтанных мутаций, почти невидимые при отборе у бактерии с множеством хромосом. Генетика 204 , 1225–1238 (2016).
CAS Статья Google ученый
Wloch, D. M., Szafraniec, K., Borts, R.H. и Korona, R. Прямая оценка частоты мутаций и распределения эффектов приспособленности в дрожжах Saccharomyces cerevisiae. Генетика 159 , 441–452 (2001).
CAS PubMed PubMed Central Google ученый
Джозеф, С. Б. и Холл, Д. В. Спонтанные мутации в диплоидных Saccharomyces cerevisiae: более выгодны, чем ожидалось. Генетика 168 , 1817–1825 (2004).
Артикул Google ученый
Зейл, К. и ДеВиссер, Дж. А. Оценки скорости и распределения эффектов приспособленности спонтанной мутации у Saccharomyces cerevisiae. Genetics 157 , 53–61 (2001).
CAS PubMed PubMed Central Google ученый
Дегнен, Г. Э. и Кокс, Е. С. Условный мутаторный ген в Escherichia coli: выделение, картирование и исследования эффекторов. J. Bacteriol. 117 , 477–487 (1974).
CAS PubMed PubMed Central Google ученый
Krašovec, R. et al. Пластичность скорости мутаций устойчивости к рифампицину зависит от межклеточного взаимодействия Escherichia coli. Nat. Commun. 5 , 3742 (2014).
ADS Статья Google ученый
Чу, X.-L. и другие. Температурные реакции частоты мутаций и спектра мутаций в штамме Escherichia coli и корреляция со скоростью метаболизма. BMC Evol. Биол. 18 , 126 (2018).
Артикул Google ученый
Shewaramani, S. et al. Escherichia coli, выращенная в анаэробных условиях, имеет повышенную частоту мутаций и четкие спектры мутаций. PLOS Genet. 13 , e1006570 (2017).
Артикул Google ученый
Булл, Дж. Дж. И Уилке, К. О. Летальный мутагенез бактерий. Генетика 180 , 1061–1070 (2008).
Артикул Google ученый
Robert, L. et al. Динамика мутаций и эффекты приспособленности отслеживались в отдельных клетках. Наука 359 , 1283–1286 (2018).
ADS CAS Статья Google ученый
Couce, A. et al. В долгосрочном эксперименте с бактериями геномы мутаторов распадаются, несмотря на устойчивый прирост физической формы. Proc. Natl Acad. Sci. США 114 , E9026 – E9035 (2017).
CAS Статья Google ученый
Baba, T. et al. Конструирование мутантов с нокаутом одного гена Escherichia coli K-12 в рамке считывания: коллекция Keio. Мол. Syst. Биол. 2 , 0008 (2006).
Артикул Google ученый
Jacquier, H. et al. Захват мутационного ландшафта бета-лактамазы ТЕМ-1. PNAS 110 , 13067–13072 (2013).
ADS CAS Статья Google ученый
Чжу, Ю. О., Сигал, М. Л., Холл, Д. В. и Петров, Д. А. Точные оценки скорости и спектра мутаций у дрожжей. PNAS 111 , E2310 – E2318 (2014).
ADS CAS Статья Google ученый
Лайонс, Д. М. и Лоринг, А. С. Доказательства избирательной основы смещения замены перехода к трансверсии в двух РНК-вирусах. Мол. Биол. Evol. 34 , 3205–3215 (2017).
CAS Статья Google ученый
Wakeley, J. Избыток переходов среди нуклеотидных замен: новые методы оценки смещения перехода подчеркивают его важность. Trends Ecol. Evol. 11 , 158–162 (1996).
CAS Статья Google ученый
Чжан, Дж. Нормы консервативных и радикальных несинонимичных нуклеотидных замен в ядерных генах млекопитающих. J. Mol. Evol. 50 , 56–68 (2000).
ADS CAS Статья Google ученый
Wan, X.-F., Xu, D., Kleinhofs, A. & Zhou, J. Количественная взаимосвязь между систематической ошибкой использования синонимичных кодонов и составом GC в одноклеточных геномах. BMC Evolut. Биол. 4 , 19 (2004).
Артикул Google ученый
Grantham, R. Формула разности аминокислот для объяснения эволюции белков. Наука 185 , 862–864 (1974).
ADS CAS Статья Google ученый
Хеникофф, С. & Хеникофф, Дж. Г. Матрицы замещения аминокислот из белковых блоков. Proc. Natl Acad. Sci. США 89 , 10915–10919 (1992).
ADS CAS Статья Google ученый
Мандал, Р. К., Цзян, Т.И Квон, Ю. М. Основной геном Campylobacter jejuni. BMC Genom. 18 , 616 (2017).
Elez, M., Radman, M. & Matic, I. Частота и структура рекомбинантных продуктов определяется клеточным уровнем MutL. PNAS 104 , 8935–8940 (2007).
ADS CAS Статья Google ученый
Hershberg, R. & Petrov, D. A. Доказательства того, что мутация универсально смещена в сторону AT у бактерий. PLOS Genet. 6 , e1001115 (2010).
Артикул Google ученый
Хильдебранд, Ф., Мейер, А. и Эйр-Уокер, А. Доказательства отбора по геномному содержанию GC в бактериях. PLOS Genet. 6 , e1001107 (2010).
Артикул Google ученый
Маккатчеон, Дж. П. и Моран, Н. А. Чрезвычайное сокращение генома у симбиотических бактерий. Nat. Rev. Microbiol. 10 , 13–26 (2011).
Артикул Google ученый
Shaver, A.C. & Sniegowski, P.D. Спонтанно возникающие мутаторы mutL в эволюционирующих популяциях Escherichia coli являются результатом изменений в длине повторов. J. Bacteriol. 185 , 6076–6082 (2003).
CAS Статья Google ученый
Ленски, Р. Э., Роуз, М. Р., Симпсон, С. К. и Тадлер, С. С. Долгосрочная экспериментальная эволюция кишечной палочки. я. адаптация и дивергенция в течение 2000 поколений. Am. Nat. 138 , 1315–1341 (1991).
Артикул Google ученый
Chevin, L.-M. Об измерении отбора в экспериментальной эволюции. Biol. Lett. 7 , 210–213 (2011).
Артикул Google ученый
Core Team, R. R: Язык и среда для статистических вычислений [Интернет] 2015 (Фонд R для статистических вычислений, Вена, Австрия, 2015).
Google ученый
Vallenet, D. et al. MaGe: система аннотации микробного генома, поддерживаемая результатами синтении. Nucleic Acids Res. 34 , 53–65 (2006).
CAS Статья Google ученый
Кавасима, С. & Канехиса, М. AAindex: база данных аминокислотных индексов. Nucleic Acids Res. 28 , 374 (2000).
CAS Статья Google ученый
Использование частот GC, кодонов и аминокислот для понимания эволюционных сил в геномном масштабе
1 Abstract
Хорошо известно, что содержание GC сильно различается между организмами; Считается, что это вызвано сочетанием мутационных предпочтений и давления отбора.Внутри кодирующих областей вариация GC более существенна в третьей позиции и меньше в первой и второй. Менее известно, что это изменение также оказывает огромное влияние на частоту аминокислот, поскольку их кодоны различаются по содержанию GC. Например, доля аланинов в разных протеомах колеблется от 1,1% до 16,5%. В общем, частота различных аминокислот сильно коррелирует с количеством кодонов, содержанием GC этих кодонов и содержанием GC генома. Однако есть явные и систематические отклонения от ожидаемых частот.Некоторые аминокислоты встречаются чаще, чем ожидалось случайно, а другие встречаются реже. Правдоподобная модель, объясняющая это, состоит в том, что на гены действуют две разные селективные силы; Во-первых, существует сила, действующая для поддержания общего уровня GC, а во-вторых, существует селективная сила, действующая на уровень аминокислот. Здесь мы используем отклонение частоты встречаемости аминокислот от ожидаемого по содержанию GC для анализа давления отбора, действующего на частоты кодонов в трех царствах жизни.Мы находим четыре основных селективных силы; Во-первых, частота серина ниже, чем ожидалось, во всех геномах, но больше всего у прокариот. Во-вторых, существует селективное давление, действующее для уравновешивания положительно и отрицательно заряженных аминокислот, что приводит к снижению содержания аргинина и отрицательно заряженных аминокислот. Это приводит к уменьшению количества аргинина и всех отрицательно заряженных аминокислот. В-третьих, частота гидрофобных остатков, кодируемых T во втором положении кодона, не изменяется с GC.Их частота у эукариот ниже, чем у прокариот. Наконец, некоторые аминокислоты с уникальными свойствами, такие как пролин глицин и пролин, имеют ограниченную вариацию частоты.
2 Введение
Частота GC значительно варьируется между разными геномами и внутри геномов, как в кодирующих, так и в некодирующих областях [1]. Причина этого не совсем понятна. Однако это, вероятно, связано с сочетанием баланса между мутационными предпочтениями, давлением отбора и историей эволюции [1].В целом мутационные предпочтения снижают уровни GC у большинства организмов [2, 3], в то время как GC-biased генная конверсия (gBGC) может способствовать более высоким уровням GC [4]. Окружающая среда организма также может влиять на уровень GC, поскольку уровни GC выше у термофилов [5]. Кроме того, существует филогенетический сигнал, так что близкородственные организмы в большинстве случаев имеют сходные уровни GC [6]. Наконец, различия в субъединице III ДНК-полимеразы могут коррелировать с различиями в GC [7].
Здесь мы не пытаемся ответить на давно обсуждаемое происхождение различий в содержании GC.Вместо этого мы предполагаем, что существует некий механизм, приводящий содержание GC в конкретном организме к оптимальному уровню. После этого мы спрашиваем, как это влияет на протеомы, исследуя частоту аминокислот, нуклеотидов и кодонов. Разное количество кодонов кодирует разные аминокислоты. Эти кодоны различаются по содержанию GC. Следовательно, в целом, аминокислоты, кодируемые большим количеством кодонов, встречаются чаще, а аминокислоты, кодируемые кодонами, богатыми GC, чаще встречаются в геномах, богатых GC [8–10], что приводит к огромным вариациям в частоте встречаемости аминокислот у разных организмов. [11, 12].Например, положительно заряженные аминокислоты Arg и Lys варьируются по частоте от 2% до 10%. Arg чаще встречается в организме с высоким содержанием GC, а Lys чаще встречается в организмах с низким содержанием GC.
Таблица кодонов, см. Рисунок 1, на удивление хорошо сохранилась с раннего детства. Один и тот же 61 кодон кодирует двадцать аминокислот у большинства организмов. Однако существуют некоторые вариации. Например, у эукариот один из стоп-кодонов может кодировать селен метионин [13], а среди Mycoplasma, Spiroplasma, Ureaplasma и Mesoplasma существуют другие варианты [14].Избыточность в таблице кодонов означает, что для многих аминокислот третье положение не меняет аминокислоту. Следовательно, общее содержание GC может изменяться при использовании разных нуклеотидов в третьем положении, не влияя на протеом. Кроме того, кодоны эволюционировали таким образом, что общие свойства аминокислот определяются главным образом кодоном во втором положении [15].
Рисунок 1.Таблицы кодонов с аминокислотами, закодированными в соответствии с различными свойствами. (a) Цвет основан на типе аминокислоты (гидрофобная — желтый, основной — синий, кислотный — красный, полярный — зеленый, амфипатический — пурпурный и коричневый с предпочтением петли) (b) окрашен в соответствии со значениями pI, чтобы быть нейтральным (c) окрашен в соответствии с предпочтением вторичной структуры и (d) окрашен в соответствии с предпочтением беспорядка.Рисунок основан на рисунке из Википедии на http://www.wikipedia.org/
Помимо частоты кодонов и уровня GC, существует множество факторов, которые влияют на частоту аминокислот [16–18]. Стоимость синтеза аминокислот может влиять на их частоту [19, 20], а некоторые аминокислоты, такие как серин, могут быть токсичными на высоких уровнях [21]. В дополнение к очищающим эффектам для снижения частоты встречаемости одной аминокислоты, организму может потребоваться минимальная частота аминокислот с определенными свойствами, в то время как другие аминокислоты, такие как аланин, могут варьироваться более свободно [22].
Ниже мы анализируем частоту аминокислот, кодонов и нуклеотидов в разных геномах. Мы показываем, что частота кодонов не полностью объясняет частоты аминокислот, то есть другие факторы также влияют на частоты аминокислот. Некоторые аминокислоты, такие как серин, встречаются реже, чем ожидалось, в то время как другие, такие как глутамат, встречаются чаще. Кроме того, некоторые аминокислоты, такие как пролин, в меньшей степени зависят от GC, чем ожидалось, что указывает на то, что существуют пределы того, насколько они могут варьироваться.Возникающая картина заключается в том, что с точки зрения генома существуют две селективные силы: одна, которая регулирует содержание GC до определенного уровня, а другая, которая при определенном уровне GC регулирует частоты аминокислот. Путем детального анализа мы можем получить представление о силах, действующих на аминокислоты.
3 Материал и методы
3.1 Наборы данных
Набор данных, используемый в этом исследовании, основан на полных бактериальных, архейных и эукариотических протеомах в UniProt [23] по состоянию на декабрь 2017 года.Все геномы из Mycoplasma, Spiroplasma, Ureaplasma и Mesoplasma были проигнорированы, поскольку в них используется другой кодон, который влияет на ожидаемую частоту аминокислот. Окончательный набор данных содержит 36 098 162 белковых последовательностей из 8 546 геномов, разделенных на 7 197 бактериальных, 351 архейных и 998 эукариотических видов. Для каждого генома содержание генома GC и длина были получены из NCBI. Далее были загружены ДНК и аминокислотные последовательности каждого гена. Обработанные наборы данных, а также все скрипты доступны в этом репозитории [24].
3.2 Статистика
Для каждого белка мы рассчитали частоты аминокислот, GC, кодонов и нуклеотидов. Средняя, максимальная и минимальная частоты для каждого генома в наборе данных представлены в таблице S1.
Таблица S1.Сводная таблица всех особенностей трех королевств. Распечатываются среднее значение, стандартное отклонение, максимальное и минимальное значения, а также коэффициент корреляции с GC-кодированием.
ANOVA типа 2 F-тесты [25] использовались для определения вкладов различий между царствами, компенсируя различия в содержании GC, см. Таблицу ?? .Используя каждый кодон / аминокислоту / нуклеотид в качестве зависимой переменной, а содержание GC использовали в качестве независимой переменной, проверяли разницу между царствами. Здесь следует отметить, что даже незначительные различия статистически значимы, поскольку набор данных велик. Кроме того, различия между эукариотами и бактериями доминируют в тесте ANOVA, так как это наиболее заметные группы.
3.3 Ожидаемые частоты
Необходимо определить ожидаемую частоту встречаемости аминокислоты ( AA i ) для определения любого селективного давления.Поэтому мы определяем модели для оценки ожидаемой частоты встречаемости аминокислот при различных сценариях. Простейшая модель, модель кодона , предполагает, что частота встречаемости аминокислоты определяется исключительно количеством кодонов, кодирующих эту аминокислоту: где Codons i — это количество кодонов для аминокислоты i , а 61 — количество кодонов, исключая стоп-кодоны.
Альтернативно, частота аминокислот может зависеть от GC (т.е.е. существует другой механизм, который определяет содержание GC в геноме), приводящий к ожидаемой частоте встречаемости аминокислот на определенном уровне GC: где Кодоны i представляют собой кодоны для аминокислоты i и x трех нуклеотидов в этом кодоне, GC — это фракция GC в геноме и δ ( N ( x ) ∈ ( A, T )) — дельта-функция, которая равна единице, если нуклеотид N ( x ) является A или T, и нулю, если нет.Кроме того, это ожидаемая частота стоп-кодонов для данных GC, как определено здесь: где ∑ стоп-кодонов суммируется по трем стоп-кодонам.
Однако, как мы покажем ниже, есть и другие параметры, которые также влияют на частоты аминокислот. Чтобы принять во внимание несколько сценариев, мы используем следующие формулы для оценки частоты встречаемости аминокислот ( AA i ) для аминокислоты i на заданном уровне GC :
Здесь — ожидаемая частота аминокислот и в GC, как определено в уравнении 2.- ожидаемая частота при GC = 50%, а W i и K i — два параметра, оптимизированные для каждой аминокислоты. Причина использования этой функции, а не просто AA i = w i * GC + k i состоит в том, чтобы иметь согласованное определение параметров W i и K i .В частности, параметр K i полезен для оценки избыточного или недостаточного представления аминокислоты.
Используя уравнение 4, мы можем моделировать различные сценарии. Если W i = 0 и K i = 0, то уравнение 4 описывает ожидаемую частоту на основе количества кодонов, как в уравнении 1 (модель кодона ). Если W i = 1 и K i = 0, уравнение описывает ожидаемую частоту аминокислот на определенном уровне GC, как в уравнении 2.Если W i = 0, а K i оптимизировано, это описывает среднюю аминокислотную частоту во всех геномах, а затем K i представляет собой сдвиг от ожидаемой частоты . Наконец, мы можем оптимизировать как W i , так и K i и получить уровни аминокислот, используя два параметра (модель twopar ). Здесь для большей реалистичности мы ограничиваем W i значениями от 0 до 1.Также здесь K i представляет отклонение от ожидаемой частоты.
Чтобы сравнить разные модели для оценки аминокислот, мы используем коэффициент корреляции Пирсона [26] и среднюю ошибку между оцененной и наблюдаемой частотами всех двадцати аминокислот, см. Рисунок S1.
Рисунок S1.Корреляция между ожидаемой и наблюдаемой аминокислотной частотой для всех аминокислот по всем геномам.
3.4 Линейные регрессии
Чтобы оценить частоту GC по частоте аминокислот, мы использовали sklearn [27].Учитывая аминокислотную частоту одной или нескольких аминокислот в белке или протеоме, модель была обучена предсказывать уровень GC кодирующей области протеома. Кроме того, мы обучили ту же модель предсказанию уровня GC для одного белка. Здесь использовалось 25000 случайно выбранных белков.
4 Результаты и обсуждение
Частота GC может сильно различаться между организмами, см. Рис. 2. В нашем наборе протеомов бета-протеобактерии Candidatus Zinderia Insecticola имеют самое низкое содержание GC — 13.5% и Geodermatophilus nigrescens имеет самый высокий (75,9%), см. Таблицу S1. Механизм, вызывающий эти различия, полностью не известен, но такие факторы, как частота мутаций, скорость кроссовера, термодинамическая стабильность и филогенетическая память, вносят свой вклад [4]. Во всяком случае, в этом исследовании мы не будем заострять внимание на различии GC. Вместо этого мы проанализируем, как разница в уровнях GC влияет на протеомы, и будем использовать отклонение от ожидаемых частот для анализа селективного давления на уровне протеома.
Рис. 2.Распределение содержания GC в трех различных царствах. На (а) содержание GC во всем геноме нанесено на график в зависимости от содержания GC в кодирующих областях. В (б-г). содержание GC в трех положениях кодона нанесено на график относительно GC в кодирующих областях.
4.1 Распределение GC
Во-первых, несколько замечаний об общем содержании GC. Оба прокариотических царства имеют бимодальное распределение GC с одним пиком около 40%, а вторым — около 70% [28], см. Рисунок 2a.Напротив, у эукариот есть единственный, менее широкий пик содержания GC. Следовательно, стандартное отклонение у прокариот больше (12% против 8%), тогда как средние уровни GC аналогичны (48% -51% для кодирующих областей), см. Таблицу S1.
4.1.1 GC-кодирование и некодирование
Для обоих прокариот геномный уровень GC (от NCBI) и уровень GC кодирующих областей (от Uniprot) почти идентичны и полностью коррелированы (CC = 0,998), тогда как для эукариот уровни немного отличаются, но все еще сильно коррелированы (CC = 0.89), см. Таблицу S1. У эукариот более высокое содержание GC в кодирующих областях (49% против 44%), см. Рисунок 2a. Уровень GC в кодирующих областях аналогичен среднему уровню, наблюдаемому у прокариот. Поэтому мы считаем, что сравнение функций с GC-содержимым кодирующей области более уместно. Кроме того, это также упрощает анализ частот кодонов и нуклеотидов.
4.2 Селективное давление на уровне GC
В таблице кодонов семь (Phe, Leu, Val, Pro, Thr, Ala и Gly) из двадцати аминокислот определены положениями один и два, см. Рисунок 1.Кроме того, два первых основания и комбинация TC или AG в третьем положении определяют восемь других аминокислот (Tyr, His, Gln, Asn, Lys, Asp, Glu и Cys). Две аминокислоты (Met и Trp) имеют только один кодон, а Ile использует три кодона ATX, не кодирующие Met. Остальные две аминокислоты, серин и аргинин, кодируются двумя группами кодонов с разными нуклеотидами в первом и втором положении. Наконец, есть три стоп-кодона, каждый из которых имеет букву T в первой позиции (T1).
Учитывая положение в таблице кодонов для аминокислот со сходными свойствами, ясно, что, в частности, положение два определяет свойства аминокислоты [15].Например, все кодоны с T2 кодируют гидрофобные аминокислоты, в то время как обе отрицательно заряженные аминокислоты имеют A2.
4.2.1 GC в разных положениях
Содержание GC различается в трех положениях кодона, см. Рисунок 2 и таблицу S1. Во всех положениях содержание GC сильно коррелирует с общим содержанием GC ( Cc > 0,93). Среднее содержание GC ниже во второй позиции, чем в двух других. Кроме того, в положениях один и два изменение содержания GC гораздо более ограничено, чем в положении три.Наивысший уровень GC на третьей позиции составляет 97%, а самый низкий — 3% по сравнению с 18% и 58% на второй позиции. Разница между позициями означает, что вариация GC в позиции 3 значительно выше, чем в других частях генома.
Модель для объяснения вариации GC в трех положениях может быть сформулирована следующим образом: В организме существует селективное давление, чтобы иметь определенное оптимальное содержание GC (в кодирующих областях). Для некоторых организмов этот оптимальный уровень очень высокий или очень низкий, т.е.е. крайний. Однако давление отбора, действующее на частоты аминокислот, делает невозможным получение экстремальных уровней GC в положениях один и два. Следовательно, чтобы получить экстремальные общие уровни GC в этих организмах, необходимо выполнить чрезмерную компенсацию в третьей позиции. Теоретически, если содержание GC ограничивается 50% в положении 1 и 2, но изменяется в положении 3, это позволяет геномному GC варьироваться от 33 до 67%. Однако содержание GC в одном и двух также варьируется, и частота аминокислот также изменяется; поэтому общее содержание ГХ может варьироваться от 13% до 76%.
Хотя среднее содержание GC одинаково во всех трех положениях, ясно, что частоты нуклеотидов не совпадают, см. Рис. 3, 4 и таблицу S1. Наибольшие различия наблюдаются в первой и второй позиции. В первом положении G1 и A1 встречаются чаще, чем два других нуклеотида, в то время как во втором положении чаще всего встречаются A2 и T2. Для более подробного понимания этих различий мы проанализируем частоты каждого нуклеотида в каждой позиции, начиная с позиции один.
Рисунок 3.Средний состав нуклеотидов в различных положениях кодонов.
Рисунок 4.Позиционные специфические частоты нуклеотидов в зависимости от частоты GC.
4.2.2 Позиция 1
В первой позиции видно, что G1 встречается наиболее часто, а T1 — наименее часто (средняя частота составляет 16,7%). Однако следует помнить, что все три стоп-кодона имеют T1, поэтому ожидаемая частота T1 составляет не 25%, а только 20,3%. Кроме того, серин, который имеет четыре из шести кодонов с T1, является одной из наиболее недопредставленных аминокислот, как мы описывали ранее [12].G1 кодирует VADEG, все эти аминокислоты избыточно представлены по сравнению со случайными, см. Рисунок 5.
Рисунок 5.Частота различных по сравнению с GC аминокислот генома сортируется по GC-содержанию кодонов. Аминокислоты отсортированы по их баллам TOP-IDP. Число представляет долю GC среди кодонов. Синяя линия представляет ожидаемую долю согласно частоте кодонов. Фиолетовые линии представляют ожидаемую долю содержания GC в положении кодона.
4.2.3 Позиция 2
Позиция 2 является наиболее консервативной позицией, когда дело касается содержания ГХ. Также ясно, что A2 и T2 встречаются чаще, чем G2 и C2, в среднем около 30% против 20%, см. Таблицу S1 и Рисунок 3. Кроме того, T2 почти не зависит от содержания GC во всех геномах, но постоянно ниже в эукариот, чем прокариот, см. рисунок 4. Постоянный уровень Т2 гарантирует стабильное количество неполярных и β -образующих аминокислот (FLIVM). G2 встречается редко и имеет ограниченный диапазон.G2 кодирует несколько аминокислот с уникальными свойствами, таких как глицин (наименьшая аминокислота) и цистеин (который может образовывать дисульфидные связи), а также аргинин, триптофан и один из стоп-кодонов. Редкость может быть обусловлена 40% (6 из 15) кодонов non-stop G2, которые кодируют аргинин, а частота аргинина недостаточно представлена, см. Рисунок 5. Наконец, A2, который кодирует в основном заряженные и полярные аминокислоты. (YHQNEDK) и C2, который кодирует APST, могут изменяться более свободно, чем два других нуклеотида в положении 2.
4.2.4 Позиция 3
В целом считается, что позиция 3 в кодоне не находится под селективным давлением, так как она редко влияет на аминокислоту, рис. 1. Однако, если селективное давление не действует на позицию три, случайный дрейф сделал бы все нуклеотиды одинаково часто в этом положении, и ясно, что это не так, см. рис. 4. Напротив, нуклеотиды в третьем положении различаются гораздо больше, чем в других положениях. Частота большинства нуклеотидов колеблется от 1% до 60%, подтверждая идею о том, что предпочтение геномной GC определяет частоты нуклеотидов.C3 наиболее часто встречается в позиции 3, но реже всего в двух других положениях, см. Рисунок 3.
4.3 Частота аминокислот по сравнению с GC
Чтобы иметь возможность идентифицировать селективное давление, действующее на частоты аминокислот, необходимо оценить ожидаемые частоты аминокислот без какого-либо давления отбора. Следовательно, необходимо иметь модель для описания ожидаемой частоты встречаемости аминокислот в геноме. Следуя приведенным выше предположениям, мы действительно предполагаем, что: существует некоторый эволюционный процесс, который стремится к адаптации содержания GC в геноме, но он не зависит от давления отбора, действующего на аминокислотном уровне.Затем можно смоделировать ожидаемые частоты аминокислот, предполагая, что области, кодирующие белок, будут случайными в отсутствие какого-либо селективного давления на уровне аминокислот.
Простым объяснением вариации частоты встречаемости аминокислот было бы то, что это просто определяется количеством кодонов, кодирующих аминокислоту, как в уравнении 1. Тем не менее, лучшее согласие наблюдается при учете GC и вычислении ожидаемые частоты встречаемости аминокислот, учитывая GC генома, как в уравнении 2.Ниже мы используем это уравнение для оценки ожидаемой частоты встречаемости аминокислот.
На рис. 5 показаны частоты встречаемости аминокислот каждой аминокислоты в сравнении с содержанием GC в кодирующих областях с синими линиями, представляющими ожидаемые частоты встречаемости аминокислот согласно уравнению 2. Сортировка аминокислот основана на содержании GC в их кодоны.
4.3.1 Частота встречаемости аминокислот с низким GC сильно зависит от GC
Частота всех аминокислот с менее чем одной третью GC в их кодонах, т.е.е. Ile, Lys, Asn, Phe и Tyr демонстрируют сильную корреляцию с GC, см. Верхний ряд на рисунке 5. Частота этих аминокислот варьируется от 1-2% при высоком GC до 19% при низком GC и корреляция с GC составляет от 0,83 до 0,93, см. таблицу S1. Наименьшие корреляции с GC имеют Tyr и Phe, которые имеют более плоское распределение, чем ожидалось, исходя только из частоты GC.
4.3.2 Частоты аминокислот с низкой зависимостью от GC не зависят от GC
Далее, существует 11 аминокислот с содержанием GC в их кодонах от одной до двух третей.Ни один из них не показывает сильной зависимости от GC, но корреляции с GC довольно высоки для валина (CC = 0,72) и Trp (CC = 0,74). В частности, некоторые из этих аминокислот встречаются чаще, чем ожидалось, из кодонов, а некоторые — реже.
4.3.3. Частота всех кодонов с высоким GC сильно зависит от GC
Наконец, аминокислоты с более чем двумя третями GC в их кодонах также сильно зависят от содержания GC (CC > 0,85). Сдвиги можно увидеть, поскольку Arg встречается реже, чем ожидалось.Частоты Gly и Pro также, похоже, ограничены определенным диапазоном.
4.4 Систематические сдвиги
Из приведенных выше исследований ясно, что существуют систематические расхождения в частотах аминокислот для некоторых аминокислот. В общем, расхождения представляют собой (комбинацию) двух типов: сдвиги и уменьшенную зависимость от GC. Сдвиг относится к тому, что частота встречаемости аминокислот постоянно чрезмерно или недостаточно представлена (как для серина), в то время как уменьшенная зависимость GC относится к уменьшенной зависимости GC, т.е.е. более плоское распределение (как для пролина), см. рисунок 5. Возникает соблазн предположить, что сдвиг указывает на то, что существует селективное давление для этой аминокислоты, которое будет более или менее частым, в то время как уменьшенная зависимость от GC указывает на то, что существует селективное давление для поддержания постоянного уровня этой аминокислоты.
Для определения систематических сдвигов мы использовали уравнение 4 (с различными ограничениями на параметры). Здесь K описывает сдвиг вверх или вниз от ожидаемого случайным образом, а W описывает силу зависимости с GC (один идеально коррелирован, а ноль указывает на отсутствие зависимости).Параметр W , следовательно, актуален только для аминокислот с GC-богатыми или GC-бедными кодонами, см. Рисунок S3.
Рисунок S2.В третьей позиции идеальная корреляция, т.е. GC определяет все.
Рисунок S3.Комбинированные графики от параметров подгонки и абсолютных ошибок.
Рисунок 6 показывает, что сдвиги ( K ) согласованы независимо от того, какая модель используется. Аргинин, серин, цистеин и пролин недостаточно представлены, в то время как глутамат, аспартат, лизин и аланин представлены чрезмерно, см. Рисунок 6.Эти сдвиги также ясно наблюдаются на Рисунке 5. Различия между царствами невелики, но сдвиги в целом меньше у эукариот. Средняя ошибка модели GC, уравнение 2, составляет 1,4% у эукариот против 1,9% у бактерий и 2,1% у архей.
Рисунок 6.Комбинированные частотные графики
4.5 Сложный баланс заряженных остатков
Положительно заряженные аминокислоты Lys и Arg похожи на сиамских близнецов, один имеет кодоны с богатым GC, а другой с низким уровнем GC, оба заряжены положительно, и они могут часто (но не всегда) выполнять аналогичные роли в белке.Одно заметное отличие состоит в том, что шесть кодонов кодируют аргинин по сравнению с двумя для лизина, то есть аргинин должен быть в три раза чаще при 50% GC. Однако аргинин постоянно встречается реже, чем ожидалось от GC, в то время как лизин встречается чаще, что компенсирует разницу в кодонах, см. Рис. 5 и 6. Общее количество Arg + Lys довольно постоянно, но немного уменьшается с GC, см. Рис. 7.
Рисунок 7.Частоты групп аминокислот по сравнению с GC.
Отрицательные аминокислоты (Asp и Glu), напротив, не сильно зависят от GC и являются постоянными в GC, см. Рисунок 5.Примечательно, что как группа отрицательно заряженные аминокислоты встречаются гораздо чаще по сравнению с тем, что ожидается случайным образом, см. Рис. 7 и 6. Таким образом, сдвиги, скорее всего, являются следствием того, что существует восемь кодонов для положительно заряженных аминокислот по сравнению с только четыре для отрицательно заряженных аминокислот и что общий заряд протеома близок к нейтральному независимо от содержания GC, см. рис. 7.
4.6 Ограниченные диапазоны частот
В дополнение к аминокислотам, которые постоянно чрезмерно или недостаточно представлены, есть существуют аминокислоты, вариации которых ограничены.На рис. 5 и S3 видно, что пять аминокислот менее зависимы от GC, чем ожидалось. Изолейцин, тирозин и фенилаланин встречаются реже, чем ожидалось, при низком уровне GC и чаще при высоком уровне GC. Точно так же Pro и Gly встречаются чаще, чем ожидалось, при низком уровне сборки мусора и реже при высоком уровне сборки мусора. Учитывая уникальные свойства Tyr / Phe (ароматический) и Gly / Pro (разрушители вторичной структуры), неудивительно, что существуют границы для их частотных вариаций.
4.7 Различия между царствами
Хотя большинство аминокислот и частот кодонов сходны в трех разных царствах, следует отметить некоторые различия. Ранее мы сообщали, что серин и пролин чаще встречаются у эукариот, а изолейцин — реже [12]. Здесь мы подтверждаем, что эти различия являются одними из наиболее значимых различий между царствами, используя тест ANOVA, см. Таблицу 1. Однако могут быть обнаружены и другие различия.
Таблица 1.ANOVA для сравнения царств. Здесь перечислены наиболее важные особенности при сравнении всех трех царств, для всех остальных сравнений см. Дополнительную таблицу ?? . Показаны средние частоты в трех царствах. Обратите внимание, что указанная здесь средняя характеристика не компенсирует различия в содержании GC, как это сделано в тесте ANOVA.
В таблице 1 можно увидеть, что две особенности доминируют в различии между царствами: повышенная частота серина у эукариот и пониженная частота T2 у эукариот.Как упоминалось выше, T2 кодирует гидрофобные аминокислоты фенилаланин, лейцин, изолейцин, метионин и валин.
Если игнорировать различия в кодонах, следующим по важности является увеличение частоты пролина в эукариотах и уменьшение частоты изолейцина [12]. Затем следует снижение уровня глицина и повышение уровня цистеина у эукариот. Наконец, G1 реже встречается у эукариот, чем у прокариот. G1 кодирует валин, аланин, аспартат, глутамат и глицин, и все они немного реже встречаются у эукариот, чем у прокариот.
Все кодоны, получившие наивысший рейтинг в тесте ANOVA, кодируют одну из обсуждаемых выше аминокислот. Интересно отметить, что кодон CCA объясняет большую часть увеличения пролина.
4.7.1 Археи
По многим характеристикам, таким как частота глутамата и аспартата, можно видеть, что царство архей разделено на две группы. Краткий анализ показывает, что это примерно коррелирует с филумом Euryarchaeota и другими археями. У евроархей больше белков (2170 против 1620), выше GC (50% против 45%) и больше Asp (6.3% против 4,9%) и Glu (8,2% против 7,2%), но меньше Lys (7,3% против 5,8%). Хотя это интересно, подробный анализ этих различий выходит за рамки целей настоящего исследования.
4.8 Прогнозирование GC по частотам аминокислот
Можно ли предсказать частоту GC по частотам аминокислот? Мы показываем, что даже частота одной аминокислоты, такой как аспарагин или аланин, в протеоме, может предсказать уровень GC с ошибкой менее 5% и коэффициентом корреляции 0,95, см. Рисунок 8.Если включить частоту всех двадцати аминокислот, ошибка упадет ниже 2%, а коэффициент корреляции составит 0,99.
Рис. 8.Прогнозирование содержания GC по частотам аминокислот.
Даже частота встречаемости аминокислот для одного белка является информативной для уровня GC всего протеома. Последовательность одного белка может предсказать уровень GC со средней ошибкой 5% и коэффициентом корреляции выше 0,90. Это можно, например, использовать для обнаружения латерально переносимых генов непосредственно из аминокислотных последовательностей, если геномная последовательность недоступна.
5 Выводы
Здесь мы изучаем взаимосвязь между содержанием GC в организмах и частотами в их кодирующих областях. Мы подчеркиваем, что частоты аминокислот значительно различаются в геномах с высоким и низким GC и что их частоты в первую очередь зависят от количества кодонов и содержания GC в их кодонах. Но есть и существенные отличия.
Чтобы объяснить это, мы предполагаем, что существует (неизвестный) механизм, действующий для поддержания уровня GC в организме.Это можно увидеть по тому факту, что третье положение различается намного больше, чем другие, и по различию частот нуклеотидов в разных положениях кодона. Далее, мы предполагаем, что существует также селективное давление, изменяющее частоты аминокислот по сравнению с ожидаемыми случайно. Этот механизм снижает частоты аргинина и серина во всех организмах, в то время как лизин, аспартат и глутамат встречаются чаще, чем ожидалось случайно. Кроме того, этот механизм ограничивает влияние GC на частоту тирозина, фенилаланина, глицина, пролина и изолейцина.
Отметим также, что селективное давление действует на; (i) Поддерживайте баланс отрицательно и положительно заряженных аминокислот во всех геномах (кроме некоторых евроархей). Это подтверждается интригой из-за недостаточного представительства аргинина и чрезмерного представительства отрицательно заряженных аминокислот. (ii) Поддержание гидрофобных остатков на постоянном уровне путем сохранения постоянной фракции тимина во втором положении кодона.
 Поиск по сайту
Поиск по сайту