
Обязательная доля в наследстве
| Помощь при Наследовании недвижимой собственности | |
|
Подготовка документов в районе наследуемого объекта недвижимости |
|
Обязательная доля в наследстве – часть наследственного имущества, независимо от содержания завещания переходящая к обязательным наследникам, которым по закону предоставлено право на обязательную долю в наследстве.
Закон определяет минимальный размер обязательной доли. Обязательная доля в наследстве не может быть уменьшена завещателем ниже определенного законом минимума.
Законодательные нормы «обязательная доля в наследстве» и «право на обязательную долю в наследстве» ограничивают свободу завещания и права завещателя.
Отказ от обязательной доли наследства
Обязательная доля наследства предназначена на поддержание несовершеннолетних и нетрудоспособных наследников или нетрудоспособных иждивенцев наследодателя, чье право на обязательную долю в наследстве и защищено законом с этой целью. Следовательно, отказываясь от обязательной доли в наследстве, обязательный наследник не имеет права отказаться в пользу других наследников, право обязательного наследника на обязательную долю не подлежит передаче другим лицам.
Оформленный у нотариуса отказ от обязательной доли наследства окончательно лишает обязательного наследника его обязательной доли в наследстве, изменить свое решение он уже будет не вправе.
Размер обязательной доли в наследстве
Обязательными наследниками наследуется обязательная доля в наследстве в размере половины доли, наследуемой каждым наследником в порядке наследования по закону, не меньше. По завещаниям, оформленным до марта 2002 года, обязательная доля будет не менее двух третей доли, наследуй они имущество по закону.
По завещаниям, оформленным до марта 2002 года, обязательная доля будет не менее двух третей доли, наследуй они имущество по закону.
Расчет обязательной доли в наследстве
происходит по определенным правилам: оценивается размер всего наследственного имущества и определяется круг наследников, которые наследовали бы данное имущество без завещания, по закону. Наследники по завещанию, не являющиеся одновременно наследниками по закону, в расчет обязательной доли не принимаются.
Порядок реализации права на обязательную долю в наследстве
Если право на обязательную долю в наследстве проигнорировано наследодателем в завещании, то по закону обязательная доля должна быть выделена обязательному наследнику. Если обязательный наследник призывается в завещании к наследованию, но завещанная ему наследодателем доля в стоимостном выражении меньше гарантированной законом обязательной доли, то недостающая часть до определенного законом размера обязательной доли должна быть компенсирована.
Право на обязательную долю в наследстве реализуется за счет других наследников.
Обязательная доля в наследстве выделяется или восполняется до необходимого размера в первую очередь из незавещанного наследственного имущества. Завещаемое имущество используется до полного осуществления права на обязательную долю, если незавещанной части имущества для этого не хватает.
Некоторые сложности могут быть при наследовании недвижимого жилого имущества или имущества, которое является источником средств существования.
Например, наследник по завещанию, в отличие от обязательного наследника, пользуется наследственным имуществом для проживания (дом, дача, квартира, другое жилье) или наследственное имущество является его основным источником средств к существованию (мастерская, орудия производства).
Если при таких условиях право обязательного наследника на обязательную долю в наследстве не может быть реализовано без лишения наследника по завещанию средств существования или жилья, то обязательная доля может быть уменьшена решением суда. Суд, учитывая все обстоятельства и имущественное положение наследников, может не только уменьшить размер обязательной доли, но и лишить обязательного наследника его права на обязательную долю.
Суд, учитывая все обстоятельства и имущественное положение наследников, может не только уменьшить размер обязательной доли, но и лишить обязательного наследника его права на обязательную долю.
Данные правила об обязательной доле и ее размере применяются с марта 2002 года. Если ныне здравствующий наследодатель составил завещание до марта 2002 года, то наследование в части права на обязательную долю будет осуществляться по старым правилам.
Различия в правилах старого и нового закона в части обязательной доли в наследстве:- размер обязательной доли в наследстве:
— по старым правилам – 2/3 полагающейся при наследовании по закону доли;
— по новым правилам – 1/2.
- уменьшение обязательной доли в наследстве судом или отказ в ее присуждении обязательному наследнику при определенных обстоятельствах:
— стало возможно по новым правилам;
— по старым правилам такого не было.
Обязательная доля в наследстве может быть учтена при наследовании на этапе ведения наследственного дела, для чего обязательный наследник должен заявить о своих наследственных правах нотариусу или решать вопрос об осуществлении своего права на обязательную долю в суде. Если свидетельство о праве на наследство нотариусом выдано наследнику (наследникам) по закону (по завещанию) до заявления обязательного наследника о своих наследственных правах, то останется только судебное решение. Обязательный наследник должен при этом понимать, что требование реализации своих наследственных прав – это не оспаривание завещания, оспаривать завещание для этого не надо.
_____________________________________________
Обязательная доля в наследстве
Обязательная доля в наследстве: кто имеет право на долю в наследстве, ее размер, можно ли оформить отказ
Право на обязательную долю в наследстве
Обязательная доля в наследстве подразумевает получение определенными гражданами наследственной массы в ситуации, когда составлено завещание, не перечисляющее их в качестве наследников.
По завещанию имущество распределяется так, как этого пожелал в своей последней воле покойный. Однако ГК РФ встает на защиту прав ряда лиц из окружения наследодателя и дает им право на наследство даже при наличии распределения имущества со стороны покойного.
Внимание, если у вас возникли вопросы вы можете их задать юристу по социальным вопросам по телефону 8 804 333 16 88 или задать свой вопрос в чате дежурному юристу. Звонки принимаются круглосуточно. Звонок бесплатный! Позвоните и решите свой вопрос!
Кто имеет обязательную долю в наследстве:
- несовершеннолетние дети;
- нетрудоспособные дети;
- ближайшие родственники – супруг, родители – при условии невозможности осуществлять трудовую деятельность;
- нетрудоспособные иждивенцы, бывшие на содержании покойного не менее года до смерти. Закон выделяет из них наследников по закону и не оговаривает необходимости совместного проживания, а также лиц, которые в число законных наследников не входят, но жили вместе с наследодателем.


Круг лиц, которые не в состоянии осуществлять трудовую деятельность, принято рассматривать в категории несовершеннолетних или нетрудоспособных
- граждане пенсионного возраста, то есть женщины от 55 и мужчины от 60 лет;
- инвалиды трех групп, а также инвалиды с детства.
В отношении обязательного наследования действуют основные моменты, касающиеся наследственного дела в целом:
Внимание, если у вас возникли вопросы вы можете их задать юристу по социальным вопросам по телефону 8 804 333 16 88 или задать свой вопрос в чате дежурному юристу. Звонки принимаются круглосуточно. Звонок бесплатный! Позвоните и решите свой вопрос!
- Обязательный наследник имеет права написать отказ от своей части. Отказ от обязательной доли в наследстве должен быть безадресным или направленным в сторону одного из наследников по завещанию. Оформление документа ведется у нотариуса, который ведет дело по наследству.
- Вступление в наследство в согласии с обязательной долей требует визита в нотариат и предоставления документов на право подобного вступления:
- паспорт;
- бумаги, которые свидетельствуют о родственной связи с покойным;
- документы для включения в круг наследников.

Что принимается:
- пенсионное свидетельство;
- справка об инвалидности;
- решение суда о недееспособности;
- признание иждивенцем: может быть получено от местной администрации на основе справок из Жилищных органов, органов социальной опеки. В ряде случаев признание факта иждивения требует обращения в судебную инстанцию и сбор доказательств. Ими могут служить платежные документы, свидетельские показания.
Размер обязательной доли в наследстве
Гражданский кодекс говорит, что обязательная часть должна составлять не меньше 50% от положенного по закону. Как это вычисляет на практике:
- Определяется вся наследственная масса и ее стоимость.
- Выявляются все законные наследники, которые при отсутствии завещания вступали бы в наследство.
- Имущество распределяется в долях. От данной доли вычисляется половина. Это и есть искомая величина.

Обязательная доля в наследстве при завещании не выступает жесткой величиной и может быть изменена. Это происходит в суде по заявлению заинтересованной стороны:
- Увеличение части. Как правило, инициируется обязательным наследником на основании того, что он фактически пользовался частью наследства, использовал его для получения дохода или осуществлял его содержание.
Пример: Екатерина является инвалидом 2 группы и входит в круг обязательных наследников своего отца. Он оставил завещание, согласно которому квартира была завещана старшему сыну, а дочь может получить другую недвижимость, принадлежавшую отцу. Девушка подает иск в суд и доказывает, что в последние 5 лет данная квартира является ее единственным местом жительства и она оплачивает ее содержание.
 Суд удовлетворяет иск и устанавливает в качестве наследства сыну передать другую жилплощадь, стоимость и размеры который не отличаются от завещанных.
Суд удовлетворяет иск и устанавливает в качестве наследства сыну передать другую жилплощадь, стоимость и размеры который не отличаются от завещанных.- Уменьшение. Инициатором выступает наследник по завещанию, часть завещанного от которого может перейти к обязательному наследнику. Основания остаются прежними.
Пример: Согласно завещанию Андрей от отца пенсионера получает его столярную мастерскую. Завещание перечисляет все имущество покойного, но не содержит сведений об обязательных наследниках. Таковыми признаются несовершеннолетние дети мужчины от второго брака. Им должна перейти доля в мастерской. Андрей в суде доказывает, что помещение является местом его работы. Суд постановляет об уменьшении обязательных долей и выделении их из иного наследуемого имущества.
Можно ли лишить наследства наследника обязательной доли?
Лишение происходит в согласии с законодательством на основании:
- Совершения наследником в отношении покойного или иных получателей наследственной массы противоправных действий, цель которых состояла в ускорении получения наследства или увеличения его массы.
- Уклонению лица от выполнения своих обязанностей по содержанию наследодателя (алименты, содержание престарелого).
- Попытка получения наследства незаконным способом (подделка документов, ложные сведения).
- Родители, которые лишены своих родительских прав.
Решение принимает суд. Поводом для возбуждения слушания становится иск от заинтересованной стороны (другие наследники).
Пример
После смерти Николая Ивановича открывается наследственное дело. Его основу составляет составленное за год до смерти завещание. В личной собственности покойного находится две квартиры общей площадью 150 квадратных метров. Они завещаны мужчиной своим детям от первого брака Ольге и Константину.
Вскоре после открытия наследства к нотариусу обращается вторая супруга покойного Александра и сообщает, что она беременна от умершего. Это подтверждается справкой из женской консультации с указанием срока беременности. Он совпадает с возможностью зачатия от наследодателя.
Женщина требует включения себя в список обязательных наследников, так как нетрудоспособна. Нотариус отказывает ей в этом, так как беременность является временной нетрудоспособностью, а для обязательной доли нужна постоянная. Но включает в список наследников нерожденного ребенка.
Наследниками по закону выступили бы двое детей первой семьи, вторая супруга и третий ребенок. Вычисляя возможные доли по закону, нотариус приходит к выводу, что нерожденный малыш наследует 18 квадратных метров в одной из квартир наследодателя.
Старшие дети предлагают Александре получить наследство в виде денежных средств за положенные метры. Женщина соглашается.
Заключение
- Наследование по завещанию включает право на обязательную часть наследства. Ей обладают нетрудоспособные и несовершеннолетние лица из числа близких родственников, а также лица на иждивении у покойного.
- Им причитается не меньше половины от доли, которую они получили бы при отсутствии последней воли. В ряде случаев размер может быть изменен судом.
- От обязательной части наследства разрешено отказаться. Также обязательного наследника можно лишить права на получение, используя нормы законодательства.
 В ряде случаев размер может быть изменен судом.
В ряде случаев размер может быть изменен судом.Наиболее популярный вопрос и ответ на него по обязательной доле в наследстве
Вопрос: Два месяца назад умер отец, он составил завещание на вторую супругу. Мы с сыном признаны обязательными наследниками. Обратились к нотариусу и узнали, что кроме квартиры у отца были большие долги. И мы их также наследуем. Можно ли этого избежать?
Ответ: Вы имеете право подать отказ от наследства. Отказаться придется от всей части: не только от наследования долгов, но и от доли квартиры. При написании отказа от лица несовершеннолетнего, каким вероятно является Ваш сын, необходимо получить разрешение от органов опеки. Рекомендуем обратиться со справкой о задолженности наследодателя, что подтверждает невыгодные условия наследования для несовершеннолетнего.

Вас так же заинтересуют следующие статьи:
Внимание, если у вас возникли вопросы вы можете их задать юристу по социальным вопросам по телефону 8 804 333 16 88 или задать свой вопрос в чате дежурному юристу. Звонки принимаются круглосуточно. Звонок бесплатный! Позвоните и решите свой вопрос!
Решение о выделении обязательной доли в наследстве
Решение о выделении обязательной доли в наследстве
РЕШЕНИЕИМЕНЕМ РОССИЙСКОЙ ФЕДЕРАЦИИ
Кузьминский районный суд г. Москвы, рассмотрев в открытом судебном заседании гражданское дело по иску о выделении обязательной доли в наследственном имуществе,
УСТАНОВИЛ:
Истица обратился в суд с вышеназванным иском, мотивируя свои требования тем, что 10.01.2011 г. ее отцом было составлено завещание на имя ответчика Михайлова Алексея, 25.01.2011 г. матерью истца было оставлено завещание на имя ответчика Михайлова Ивана.
28.04.2011 г. умерла мать истца, 07.11.2011 г. умер отец истца. В наследственную массу после смерти родителей входит квартира.
На момент открытия наследства как после смерти отца, так и после смерти матери истица являлась нетрудоспособной, в связи с чем истица полагает, что независимо от существа завещания имеет право на обязательную долю в наследстве после смерти своих родителей, просила суд выделить ей обязательную долю в наследственном имуществе в виде 1/2 доли в квартире.
Истица и ее представитель адвокат Жукова О.С. в суд явились, исковые требования поддержали, настаивали на их удовлетворении.
Представитель ответчика Михайлова А. в суд явился, иск не признал, указав, что истец не имеет права на обязательную долю в наследстве, поскольку не входит в круг лиц, определенных законом, на которых распространяется право требования обязательной доли в наследстве. Дополнительно указал, что для ответчика спорное жилое помещение является единственным местом жительства.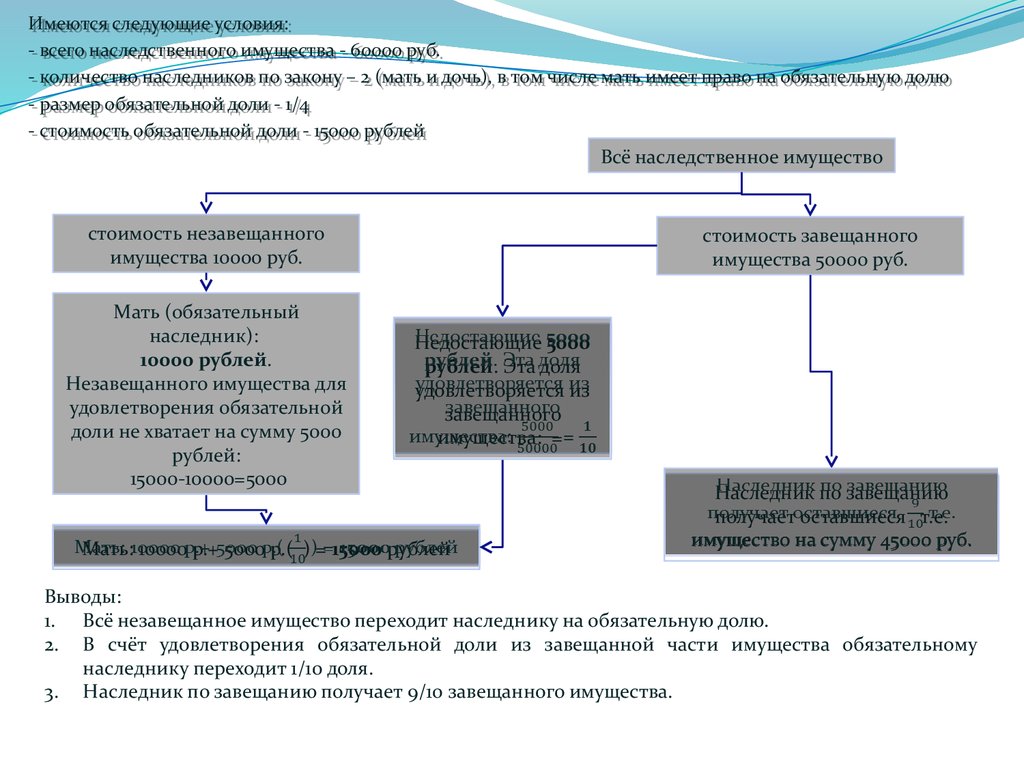
Представитель ответчика Михайлова И. в судебное заседание явился, иск не признал, указав, что истец не имеет право на обязательную долю в наследстве, поскольку на момент открытия наследства являлся трудоспособным лицом, кроме того, указал, что ответчик совместно со своей матерью и наследодателем до момента смерти проживали в спорной квартире, которая является для ответчика единственным местом жительства, в то время как истцу принадлежит по праву собственности 1/4 доля в иной квартире.
Суд, выслушав стороны, допросив свидетелей, проверив материалы дела, находит иск подлежащим удовлетворению по следующим основаниям.
В ходе рассмотрения дела установлено, что 28.04.2011 г. умерла мать истицы. При жизни умершая распорядилась своим имуществом на случай смерти путем составления 25.01.2011 г. завещания, удостоверенного нотариусом города Москвы Степновой П.Т. согласно которого все имущество, которое ко дню смерти окажется принадлежащим ей, она завещает ответчику Михайлову Ивану.
После смерти матери истца нотариусом г. Москвы Степновой П.Т. было открыто наследственное дело. Согласно материалов наследственного дела с заявлениями о принятии наследства обратились истица (дочь умершей, наследуя по закону) и ответчик Михайлов Алексей (в порядке наследования по завещанию).
На момент смерти наследодателю принадлежала по праву собственности 1/2 доля в праве собственности на квартиру (на основании договора передачи от 25.06.1993 г. и решения Кузьминского районного суда г. Москвы).
07.11.2011 г. умер отец истца. При жизни умерший распорядился своим имуществом на случай смерти путем составления 10.01.2011 г. завещания, удостоверенного нотариусом города Москвы Иваненко Т.А., согласно которого все имущество, которое ко дню смерти окажется принадлежащим ему он завещает ответчику Михайлову Алексею.
После смерти отца истицы нотариусом г. Москвы Белицкой П.Т. было открыто наследственное дело. Согласно материалов наследственного дела с заявлениями о принятии наследства обратились истица (дочь умершего, наследуя по закону) и ответчик Михайлов Алексей (в порядке наследования по завещанию).
На момент смерти наследодателю принадлежала по праву собственности 1/2 доля в праве собственности на квартиру (на основании договора передачи от 25.06.1993 г. и решения Кузьминского районного суда г. Москвы).
Свидетель Борисова, свидетель Тихонов показали суду, что неоднократно видели, как истица помогает своим родителям, носит им еду, поскольку проживала несколькими этажами выше, заботилась о родителях вплоть до смерти.
Свидетель Маленков показал, что знает истицу и ее умерших родителей, а также ответчика Михайлова Алексея. Истица у него снимала квартиру с мая 2010 года по август 2011 года, а впоследствии переехала в спорную квартиру.
Свидетель Шайко показала, что является подругой ответчика Михайлова Алексея, совместно с ним снимала квартиру, указала, что ответчик в 2011 году переехал в спорную квартиру.
Свидетель Лесков показал, что часто приходил навещать умерших бабушку и дедушку, ответчик в квартире не проживал, он просто ухаживал за ними, получая за это денежное вознаграждение.
В соответствии со ст.1110 Гражданского кодекса РФ при наследовании имущество умершего (наследство, наследственное имущество) переходит к другим лицам в порядке универсального правопреемства, то есть в неизменном виде как единое целое и в один и тот же момент, если из правил настоящего Кодекса не следует иное.
В соответствии со ст.1118 Гражданского кодекса РФ распорядиться имуществом на случай смерти можно только путем совершения завещания.
В соответствии со ст.1119 Гражданского кодекса РФ завещатель вправе по своему усмотрению завещать имущество любым лицам, любым образом определить доли наследников в наследстве, лишить наследства одного, нескольких или всех наследников по закону, не указывая причин такого лишения, а в случаях, предусмотренных настоящим Кодексом, включить в завещание иные распоряжения. Завещатель вправе отменить или изменить совершенное завещание в соответствии с правилами статьи 1130 настоящего Кодекса.
Свобода завещания ограничивается правилами об обязательной доле в наследстве (статья 1149).
В соответствии со ст.1149 Гражданского кодекса РФ несовершеннолетние или нетрудоспособные дети наследодателя, его нетрудоспособные супруг и родители, а также нетрудоспособные иждивенцы наследодателя, подлежащие призванию к наследованию на основании пунктов 1 и 2 статьи 1148 настоящего Кодекса, наследуют независимо от содержания завещания не менее половины доли, которая причиталась бы каждому из них при наследовании по закону (обязательная доля).
В соответствии со ст. 2 ФЗ «О трудовых пенсиях в РФ» трудовая пенсия – ежемесячная денежная выплата в целях компенсации застрахованным лицам заработной платы и иных выплат и вознаграждений, утраченных ими в связи с наступлением нетрудоспособности вследствие старости или инвалидности, а нетрудоспособным членам семьи застрахованных лиц – заработной платы и иных выплат и вознаграждений кормильца, утраченных в связи со смертью этих застрахованных лиц, право на которую определяется в соответствии с условиями и нормами, установленными настоящим Федеральным законом.
При этом наступление нетрудоспособности и утрата заработной платы и иных выплат и вознаграждений в таких случаях предполагаются и не требуют доказательств.
В соответствии со ст. 7 ФЗ «О трудовых пенсиях в РФ» право на трудовую пенсию по старости имеют мужчины, достигшие возраста 60 лет, и женщины, достигшие возраста 55 лет.
На момент открытия наследства после смерти матери истицы 28.04.2011 г., а также отца истицы 07.11.2011 г. истица являлась нетрудоспособной, поскольку к моменту открытия наследства достигла пенсионного возраста (1947 года рождения, на момент открытия наследства 64 года), в соответствии с удостоверением бессрочно назначена пенсия по старости.
Таким образом, доводы ответчиков об отсутствии у истицы права на обязательную долю суд считает несостоятельными.
В соответствии со ст.1049 Гражданского кодекса РФ нетрудоспособные дети наследодателя наследуют независимо от содержания завещания не менее половины доли, которая причиталась бы каждому из них при наследовании по закону (обязательная доля).
В соответствии с ч. 4 ст.1149 Гражданского кодекса РФ если осуществление права на обязательную долю в наследстве повлечет за собой невозможность передать наследнику по завещанию имущество, которым наследник, имеющий право на обязательную долю, при жизни наследодателя не пользовался, а наследник по завещанию пользовался для проживания (жилой дом, квартира, иное жилое помещение, дача и тому подобное) или использовал в качестве основного источника получения средств к существованию (орудия труда, творческая мастерская и тому подобное), суд может с учетом имущественного положения наследников, имеющих право на обязательную долю, уменьшить размер обязательной доли или отказать в ее присуждении.
Таким образом, уменьшение размера обязательной доли либо отказ в ее присуждении является правом, а не обязанностью суда. Кроме того, суд учитывает, что ответчики лишь ухаживали за родителями истицы. Истица проживает в коммунальной квартире с сыном и двумя внуками и обладает 1/4 долей в праве собственности, вследствие чего имеет существенный интерес к наследственному имуществу.
При данных обстоятельствах суд не усматривает оснований для уменьшения размера обязательной доли либо отказа в ее присуждении, вследствие чего находит исковые требования подлежащими удовлетворению и возможным выделения истицы обязательную долю в виде 1/2 доли в спорной квартире.
На основании изложенного, руководствуясь ст. 194-199 ГПК РФ, суд
РЕШИЛ:
Выделить истице обязательную долю в наследственном имуществе в виде 1/2 доли в квартире, причитающуюся ей после смерти отца и матери истицы.
Решение может быть обжаловано в Московский городской суд в течение месяца путем подачи апелляционной жалобы через Кузьминский районный суд г. Москвы.
Право на обязательную долю в наследстве
Право на обязательную долю наследства, установленное статьей 1149 Гражданского
кодекса, ограничивает последнюю волю наследодателя. Выделение такой доли
в обязательном порядке предусмотрено для некоторых лиц, которые имели бы
право на часть наследства по закону, если бы не было завещания. Они называются
Они называются
обязательными, или необходимыми, наследниками.
К ним закон относит нетрудоспособных родителей, детей (в том числе усыновленных) и
пережившего супруга. Все эти лица входят в первую очередь наследников по закону. Не
имеет значения, состояли они на иждивении завещателя или нет. Помимо перечисленных
право на обязательную долю имеют нетрудоспособные иждивенцы, для которых
поддержка умершего была единственным (или основным) источником средств на
протяжении года до его смерти.
В каких случаях необходимый наследник может воспользоваться своим правом?
- если в отношении него завещателем не сделаны никакие распоряжения;
- когда он лишен завещателем наследства;
- когда завещанная ему или полученная по закону часть имущества меньше обязательной доли.
Обязательная доля выделяется только при ее требовании. При выявлении всех
имеющихся наследников нотариус разъясняет таким лицам их права.
Правила расчета и выделения обязательной доли
Для правильного определения размера необходимой доли сначала нужно:
- выявить полный объем и стоимость всего имущества (вещи и права), формирующего состав наследства;
- рассчитать теоретический размер доли, которую получил бы необходимый наследник, если бы наследование производилось по закону, и он бы не был лишен своего права.

При расчете объема во внимание принимается полный размер всего имущества,
как завещанного, так и оставшегося без распоряжения. В него также включаются
денежные вклады наследодателя, независимо от того, сделано ли в отношении
них завещательное распоряжение. Далее, исходя из количества наследников,
рассчитывается размер обязательной доли. Например, имеются три первоочередных
наследника: нетрудоспособная мать, жена и несовершеннолетний сын. Наследодатель
оставил завещание, в котором не упомянута мать. Каждому из них по закону положена
1/3 часть наследства. И хотя приоритетом считается воля умершего завещателя, как
устанавливает наследственное право, обязательная доля матери составляет в данном
случае — 1/6 наследства.
Обязательная доля выделяется из имущества, нераспределенного по завещанию. Но
если его недостаточно, недостающая часть восполняется из завещанного имущества.
Если в составе наследства присутствуют исключительные права, например, право на
издание рукописи наследодателя, то обязательному наследнику может быть присуждена часть наследства в процентном отношении. Он вправе получать свою долю дохода от
Он вправе получать свою долю дохода от
осуществления таких прав.
Права и обязанности необходимого наследника
Необходимый наследник, имея право на обязательную долю, может ее потребовать,
но не обязан это делать. Если ему завещано какое-то имущество, в том числе
завещательный отказ, он может от него отказаться, не утрачивая при этом свое право.
Но если он получает часть наследства по закону (когда, например, не все имущество
было завещано) и принимает его, то отказаться от этой части он не вправе. Она будет
засчитана в обязательную долю. Обязательный наследник не может отказаться от своего
права в чью-то пользу и его право не может быть передано как право представления.
Необходимый наследник, принявший наследство, отвечает по всем обязательствам
наследодателя так же, как и другие. Исключение касается только завещательного
отказа. Его выполнение возможно только за счет части наследства, полученной таким
наследником сверх обязательной части доли. Так же, как и любой наследник, он может
Так же, как и любой наследник, он может
быть признан недостойным.
Нетрудоспособное лицо, имеющее право на обязательное наследство, может получить
от нотариуса соответствующее свидетельство, предоставив подтверждающие документы.
Согласия других наследников для этого не требуется. Если возникает спор, то он может
быть решен в судебном порядке.
Опыт России и зарубежных стран Александры Фокиной :: SSRN
8 страниц Опубликовано: 15 декабря 2021 г.
Дата написания: 20 ноября 2021 г.
Аннотация
В исследовании представлен сравнительно-правовой анализ института обязательной доли в наследстве в праве России и некоторых зарубежных стран по трем критериям сравнения: понятие и правовое определение; размер обязательной доли и обязательных наследников; особенности расчета размера обязательной доли.
Актуальность выбранной темы обусловлена тем, что сходства и различия, установленные в результате сравнительно-правового анализа норм наследственного права России и зарубежных государств правового регулирования института обязательной доли в наследстве позволяют нам провести переоценку действующего регулирования обязательной доли в наследстве в российском праве и использовать опыт зарубежных стран для дальнейшего совершенствования российского законодательства о наследовании.
В результате проведенного исследования установлены существенные отличия в нормативно-правовом регулировании института обязательной доли в наследстве, которые могут быть использованы в рамках реформы наследственного права России.
Ключевые слова: Наследование по закону, обязательный наследник, обязательная доля, сравнительное правоведение
Рекомендуемое цитирование: Рекомендуемая ссылка
Начало работы в сельском хозяйстве: наследование фермы
Начало работы в сельском хозяйстве: наследование фермы
Наследование фермы может быть как захватывающим, так и пугающим.Некоторые люди точно знают, что они хотели бы делать с фермой, но многие не знают. Перед принятием окончательных решений необходимо решить ряд вопросов и проблем.
Вопрос о том, что делать с унаследованными сельскохозяйственными угодьями, становится все более актуальным. Средний возраст владельцев сельскохозяйственных угодий продолжает увеличиваться. По данным Сельскохозяйственной переписи 2017 года, средний возраст основного оператора фермы составлял 57,5 лет. Самая быстрорастущая группа операторов — это фермеры старше 65 лет.Значительно увеличился и средний возраст землевладельцев. Согласно исследованию собственности и владения сельскохозяйственными угодьями в Айове, проведенному в 2017 году, 60% сельскохозяйственных угодий в Айове принадлежало людям старше 65 лет. Владельцам старше 75 лет принадлежало 34% сельскохозяйственных угодий в Айове.
В последние годы стоимость сельскохозяйственных угодий увеличилась, но в последнее время стабилизировалась. Средняя стоимость фермерской недвижимости в США в 2020 году составила 3160 долларов за акр, как и в 2019 году, но на 5,3% выше, чем пять лет назад. Сегодня на рынок сельскохозяйственных земель влияет множество факторов. Важным фактором являются цены на товары или доход, который можно получить от земли. Другими ключевыми компонентами, влияющими на стоимость земли, являются сумма долга, связанного с приобретением земли, и государственная политика, особенно политика, связанная с торговлей, энергетикой, налогами и входными затратами. Показатели экономики США и экономики во всем мире влияют на цены на сырьевые товары, которые, в свою очередь, влияют на стоимость земли. Государственная денежно-кредитная политика является важным фактором для наблюдения, поскольку она связана с инфляцией и процентными ставками.Проблемы, связанные с погодой, как здесь, так и за рубежом, продолжают оказывать влияние на стоимость земли. Разрастание городов, развитие недвижимости и другие вопросы планирования землепользования влияют на стоимость и доступность земли.
Важным фактором являются цены на товары или доход, который можно получить от земли. Другими ключевыми компонентами, влияющими на стоимость земли, являются сумма долга, связанного с приобретением земли, и государственная политика, особенно политика, связанная с торговлей, энергетикой, налогами и входными затратами. Показатели экономики США и экономики во всем мире влияют на цены на сырьевые товары, которые, в свою очередь, влияют на стоимость земли. Государственная денежно-кредитная политика является важным фактором для наблюдения, поскольку она связана с инфляцией и процентными ставками.Проблемы, связанные с погодой, как здесь, так и за рубежом, продолжают оказывать влияние на стоимость земли. Разрастание городов, развитие недвижимости и другие вопросы планирования землепользования влияют на стоимость и доступность земли.
Индивидуальные обстоятельства диктуют наиболее желательный образ действий. В некоторых случаях земля уже может обрабатываться унаследовавшим ее лицом, и решение может быть относительно простым. В других случаях землю может унаследовать тот, кто мало знает о современном сельском хозяйстве и понятия не имеет, что делать с фермой.Наконец, ферма может быть унаследована совместно, и пожелания всех сторон могут повлиять на решение о том, что делать с фермой. Независимо от обстоятельств, каждая ситуация уникальна, и важно тщательно оценить ситуацию.
В других случаях землю может унаследовать тот, кто мало знает о современном сельском хозяйстве и понятия не имеет, что делать с фермой.Наконец, ферма может быть унаследована совместно, и пожелания всех сторон могут повлиять на решение о том, что делать с фермой. Независимо от обстоятельств, каждая ситуация уникальна, и важно тщательно оценить ситуацию.
Есть два основных фактора, которые необходимо учитывать перед принятием каких-либо решений. Во-первых, важно оценить землю, которую вы унаследовали, и понять ферму. В большинстве случаев это определяет наилучший план действий.Вторым важным фактором, который следует учитывать, является то, была ли ферма унаследована одним человеком или группой и каким образом (общая или совместная аренда). Чем больше людей занимается наследством, тем сложнее процесс принятия решения.
Эта публикация предназначена для того, чтобы поднять вопросы, которые необходимо задать, и направить наследников к конкретной информации для их обстоятельств. Эта публикация является обновленной версией более ранней публикации, подготовленной как одна из региональных дополнительных публикаций Северо-Центрального региона по началу работы в сельском хозяйстве.Эта публикация неоднократно обновлялась Джоном Бейкером, Майком Даффи и другими специалистами Университета штата Айова с момента ее написания. Многие из представленных идей схожи, потому что основа для принятия правильных решений в отношении сельскохозяйственных угодий не изменилась. Подтверждение делается в общем смысле и не будет даваться повсюду, чтобы не нарушать ход публикации.
Эта публикация является обновленной версией более ранней публикации, подготовленной как одна из региональных дополнительных публикаций Северо-Центрального региона по началу работы в сельском хозяйстве.Эта публикация неоднократно обновлялась Джоном Бейкером, Майком Даффи и другими специалистами Университета штата Айова с момента ее написания. Многие из представленных идей схожи, потому что основа для принятия правильных решений в отношении сельскохозяйственных угодий не изменилась. Подтверждение делается в общем смысле и не будет даваться повсюду, чтобы не нарушать ход публикации.
Оценка земли
Местоположение
Старая поговорка об оценке недвижимости гласит, что три наиболее важных фактора, определяющих стоимость земельного участка, — это местоположение, местоположение и еще раз местоположение.Местоположение является ключом к ценности унаследованной фермы. Местоположение — это то, что определяет текущее, лучшее и будущее использование земли. Например, земля, расположенная вблизи расширяющегося мегаполиса, имеет потенциально большую ценность для развития, чем аналогичная земля, расположенная в сельской местности. Высокопродуктивная земля стоит больше, чем низкокачественная земля для выращивания сельскохозяйственных культур. Есть много других факторов, связанных с местоположением, которые определяют стоимость земли.
Например, земля, расположенная вблизи расширяющегося мегаполиса, имеет потенциально большую ценность для развития, чем аналогичная земля, расположенная в сельской местности. Высокопродуктивная земля стоит больше, чем низкокачественная земля для выращивания сельскохозяйственных культур. Есть много других факторов, связанных с местоположением, которые определяют стоимость земли.
Доход
Потенциальный доход от недвижимости является самым важным показателем стоимости после местоположения.Расчет потенциального дохода фермы поможет оценить стоимость, а также предоставит информацию, которая поможет принять окончательное решение о том, что делать с фермой.
Текущее использование земли является хорошей отправной точкой для оценки потенциального дохода от земли. Нынешнее использование земли определяется многими факторами; поэтому важно не предполагать, что нынешнее использование земли является наилучшим, особенно при новом владении.
Оценка потенциального дохода – непростая задача. Присущая почве продуктивность поможет в оценке ожидаемой урожайности различных культур и систем земледелия. Программа Extension and Outreach Университета штата Айова может предоставить расчетные производственные затраты и среднюю урожайность по округу за прошлые периоды. Служба охраны природных ресурсов Министерства сельского хозяйства США может предоставить рекомендации по наиболее подходящему использованию земли с учетом ее природных ограничений. NRCS также может помочь разработать план сохранения почвы, если это необходимо.
Присущая почве продуктивность поможет в оценке ожидаемой урожайности различных культур и систем земледелия. Программа Extension and Outreach Университета штата Айова может предоставить расчетные производственные затраты и среднюю урожайность по округу за прошлые периоды. Служба охраны природных ресурсов Министерства сельского хозяйства США может предоставить рекомендации по наиболее подходящему использованию земли с учетом ее природных ограничений. NRCS также может помочь разработать план сохранения почвы, если это необходимо.
Доход от домашнего скота может играть важную роль в определении потенциального дохода от собственности.Например, земля, которая находится на постоянном пастбище или сенокосе, будет использоваться в основном для выращивания говядины, овец или молочного скота. В этом случае доход от скота будет определять стоимость земли.
Несколько государственных и частных организаций предлагают программы, которые могут повлиять на доход от фермы в зависимости от типа фермы и ее местоположения. Агентство сельскохозяйственных услуг Министерства сельского хозяйства США может предоставить информацию о текущих государственных программах и правах фермы на участие в этих программах. Кроме того, несколько групп по сохранению ресурсов, таких как American Farmland Trust, Nature Conservancy и Ducks Unlimited, предлагают программы, которые могут повлиять на использование фермы.Аренда для отдыха и охоты популярна во многих частях страны, и такое использование может увеличить доход или стоимость земли.
Агентство сельскохозяйственных услуг Министерства сельского хозяйства США может предоставить информацию о текущих государственных программах и правах фермы на участие в этих программах. Кроме того, несколько групп по сохранению ресурсов, таких как American Farmland Trust, Nature Conservancy и Ducks Unlimited, предлагают программы, которые могут повлиять на использование фермы.Аренда для отдыха и охоты популярна во многих частях страны, и такое использование может увеличить доход или стоимость земли.
Важно знать о любых контрактах, аренде, правилах зонирования или других сервитутах, связанных с фермой. Такие договоренности ограничивают потенциальное использование имущества в краткосрочной перспективе. Если земля в настоящее время находится в аренде, она может быть недоступна до истечения срока аренды.
Стоимость
Полная оценка сертифицированным оценщиком является наилучшим способом определения стоимости фермы.Существуют и другие методы оценки стоимости недвижимости, но они не столь точны. Использовать ли полную оценку или какие-либо другие средства, зависит от желаемой степени точности. В большинстве ситуаций, в которых участвуют несколько человек, наилучшим подходом является нейтральная оценка третьей стороной.
Использовать ли полную оценку или какие-либо другие средства, зависит от желаемой степени точности. В большинстве ситуаций, в которых участвуют несколько человек, наилучшим подходом является нейтральная оценка третьей стороной.
При оценке стоимости фермы наследник должен учитывать любые постройки, сооружения и другие улучшения земли. В некоторых случаях здания могут значительно увеличить стоимость имущества и существенно повлиять на то, как оно будет использоваться.Жилье в собственности также может влиять на использование собственности. Всегда следует определять состояние зданий, жилищ и улучшений, поскольку они помогают определить относительную стоимость имущества. В некоторых случаях старые ветхие сооружения могут фактически снизить стоимость земли, поскольку их придется снести или отремонтировать.
Размер фермы также влияет на решение о том, что с ней делать. Размер определяет стоимость наследства и возможность зарабатывать на жизнь только за счет фермы.
Во многих случаях ферма будет иметь сентиментальную ценность, которая будет играть роль при принятии решения. Сентиментальную ценность гораздо сложнее измерить количественно. Сентиментальная привязанность одного наследника к земле не будет такой же, как у других.
Сентиментальную ценность гораздо сложнее измерить количественно. Сентиментальная привязанность одного наследника к земле не будет такой же, как у других.
Налоги
Налоги на имущество и наследство как на уровне штата, так и на федеральном уровне являются важными факторами. Минимизация этих налогов может увеличить стоимость наследства, но просто минимизация налогов может не дать наилучшего использования, учитывая индивидуальные обстоятельства.
Размер поместья, то, как активы находились в собственности или в аренде, отношения наследников и то, что делается с имуществом, определяют размер налогов. Если большая часть наследства приходится на землю, может потребоваться продать часть активов или занять деньги только для того, чтобы оформить имущество.
В некоторых случаях наследство может подпадать под оценку «специального использования». В этой ситуации могут быть ограничения на то, как долго собственность может быть обработана, чтобы поддерживать эту оценку специального использования квалифицированным наследником.
В некоторых особых случаях недвижимость может быть продана, а вырученные средства реинвестированы без возникновения налоговых последствий.
Если ферма удерживается в качестве инвестиции, то, кто ее обрабатывает и имеет ли место материальное участие, может иметь важное значение при определении суммы налога на наследство.
В этой публикации невозможно рассмотреть все детали и обстоятельства налогов. Наследникам сельскохозяйственных угодий следует обратиться за квалифицированной юридической консультацией в отношении наследства, чтобы определить, на какие специальные положения оно распространяется и какие существуют ограничения на использование собственности.В некоторых случаях будет лучше заплатить налоги. В других случаях будет лучше изменить использование или распоряжение фермой, чтобы уменьшить налоговое бремя.
Если есть задолженность по налогам, новый владелец может занять средства для их уплаты, используя имущество в качестве залога.
Опции для фермы
Независимо от того, наследуется ли сельскохозяйственная земля отдельным лицом или группой, у каждого наследника есть три основных варианта: обрабатывать землю, оставить землю в качестве инвестиции или продать ферму.
Обработка земли
Обработка земли — это сложное решение, требующее учета ряда соображений. Один из первых вопросов – есть ли у наследника необходимые навыки и ресурсы.
Если наследник уже занимается сельским хозяйством, то следующим шагом будет определение того, как ферма вписывается в его текущую деятельность. Наследник мог решить добавить землю к своей текущей деятельности, чтобы сформировать одну большую единицу, или он мог продать одну из ферм и объединиться вокруг другой фермы.Перед принятием решения о том, следует ли обрабатывать унаследованное имущество или объединить его с существующей операцией, необходимо решить несколько вопросов. Насколько близко унаследованное свойство к текущему? Принесет ли размер нового предприятия адекватный доход? Сможет ли наследник справиться с этим? Потребуется ли это новое или другое оборудование?
Если наследник в настоящее время не занимается сельским хозяйством, но хотел бы начать, существует множество семейных решений относительно изменения образа жизни, которое повлечет за собой сельское хозяйство. Может быть легко увлечься очарованием сельского хозяйства или пребывания на земле. Но это решение, которое требует тщательного рассмотрения.
Может быть легко увлечься очарованием сельского хозяйства или пребывания на земле. Но это решение, которое требует тщательного рассмотрения.
Пятьдесят восемь процентов ферм имеют годовой объем продаж менее 10 000 долларов США (перепись сельского хозяйства 2017 года). Такая маленькая ферма не может содержать семью. В этих случаях фермеру нужна работа вне фермы, чтобы обеспечить необходимый доход. С другой стороны, внефермерский доход может дополнить начинающего фермера, у которого есть возможность работать полный рабочий день.Необходимо решить вопрос о том, является ли фермерское хозяйство достаточно крупным и имеет ли он потенциал для поддержки наследника и его или ее семьи в том образе жизни, к которому они привыкли или к которому стремятся. Согласно сельскохозяйственной переписи 2017 года, 61% основных операторов работали вне фермы для получения дополнительного дохода, а 40% основных операторов работали вне фермы 200 и более дней.
Здоровье оператора фермы является важным фактором, который нельзя игнорировать. Есть ли у наследника возможность заниматься физическим трудом и полностью подвергаться воздействию непогоды? Многие сельскохозяйственные работы приходится выполнять в ненастную погоду.
Есть ли у наследника возможность заниматься физическим трудом и полностью подвергаться воздействию непогоды? Многие сельскохозяйственные работы приходится выполнять в ненастную погоду.
После того, как наследник решил обрабатывать землю, он или она должны решить, выращивать ли урожай, скот или и то, и другое. ISU Extension and Outreach предлагает множество публикаций, объясняющих, какие культуры можно выращивать в штате, а также оценки затрат на производство сельскохозяйственных культур. Эту информацию можно использовать для оценки потенциального дохода от унаследованной фермы.Выбор растениеводческих и животноводческих предприятий иногда дополняет друг друга. Например, одним из вариантов является выращивание сельскохозяйственных культур, которые будут скармливаться скоту.
Производственное сельское хозяйство включает в себя гораздо больше, чем просто владение землей. Он включает в себя труд, капитал и управление. Размер капитала, необходимого для производственного сельского хозяйства, зависит от выбранной системы. Сегодня многие операции в значительной степени зависят от покупных ресурсов, что требует значительных капиталовложений. Многие операторы должны занимать средства хотя бы на часть расходов.Другие операции в большей степени зависят от рабочей силы, навыков управления или маркетинга. Обсудите свою ситуацию с потенциальными кредиторами, прежде чем брать на себя окончательное обязательство.
Сегодня многие операции в значительной степени зависят от покупных ресурсов, что требует значительных капиталовложений. Многие операторы должны занимать средства хотя бы на часть расходов.Другие операции в большей степени зависят от рабочей силы, навыков управления или маркетинга. Обсудите свою ситуацию с потенциальными кредиторами, прежде чем брать на себя окончательное обязательство.
Потребность в оборудовании будет независимо от операций. Современное сельскохозяйственное оборудование может быть дорогим и требует навыков для его безопасной и эффективной эксплуатации. Однако владение техникой — это только один из вариантов. Сегодня многие фермеры используют лизинг или индивидуальный прокат как способ управления техникой. Лизинг или индивидуальная аренда особенно выгодны для дорогого, редко используемого оборудования.
Для ведения прибыльного фермерского хозяйства необходимы хорошие управленческие навыки. Управленческие навыки в области производства и операций, финансов, общего бизнеса, личных отношений и принятия решений аналогичны другим видам бизнеса. Компьютерные или технические навыки имеют жизненно важное значение для ведения учета и бизнес-анализа и эксплуатации оборудования, особенно на крупных предприятиях. Фермер должен эффективно выявлять изменения, происходящие в отрасли, и разрабатывать стратегии их использования в личных целях, развивать позитивные отношения с покупателями и продавцами, сообщать о проблемах, определять конкурентные преимущества своей деятельности с точки зрения производительности и технической эффективности, а также разрабатывать стратегии преодоления барьеры.Некоторые методы управления рисками, используемые для устранения уязвимостей в работе, такие как страхование урожая или скота, уникальны для сельского хозяйства.
Компьютерные или технические навыки имеют жизненно важное значение для ведения учета и бизнес-анализа и эксплуатации оборудования, особенно на крупных предприятиях. Фермер должен эффективно выявлять изменения, происходящие в отрасли, и разрабатывать стратегии их использования в личных целях, развивать позитивные отношения с покупателями и продавцами, сообщать о проблемах, определять конкурентные преимущества своей деятельности с точки зрения производительности и технической эффективности, а также разрабатывать стратегии преодоления барьеры.Некоторые методы управления рисками, используемые для устранения уязвимостей в работе, такие как страхование урожая или скота, уникальны для сельского хозяйства.
Решение об обработке унаследованной земли не должно приниматься легкомысленно. Основным соображением является предполагаемый доход, который может быть получен от фермы, и достаточно ли этого для наследника. Каждая ситуация отличается.
Годовой доход фермера может сильно варьироваться от года к году. Изменения цен на сырьевые товары, производственных затрат, стоимости земли, погоды и урожайности — это лишь некоторые из факторов, влияющих на доход фермера.
Изменения цен на сырьевые товары, производственных затрат, стоимости земли, погоды и урожайности — это лишь некоторые из факторов, влияющих на доход фермера.
Владение землей в качестве инвестиции
Владение землей в качестве инвестиции — еще один вариант для наследников. Те, кто решит владеть землей, могут выбрать один из нескольких различных методов управления, каждый из которых требует разного количества времени и усилий. Некоторым людям нравится быть активными в управлении фермой, а другие предпочитают не участвовать в этом.
Сдача в аренду
Сдача земли в аренду другим лицам сегодня является жизнеспособным и распространенным вариантом. По данным сельскохозяйственной переписи, в 2017 году 51% земель в фермерских хозяйствах был сдан в аренду.Возможны различные варианты аренды. Подробное обсуждение договоров аренды см. на веб-сайте ответственного за принятие решений в области сельского хозяйства Университета штата Айова. Типичный лизинговый договор включает денежную ренту, гибкую денежную арендную плату или долю урожая. В ситуации с наличными арендатор несет большую часть риска. В соглашении о разделе урожая производственный и ценовой риск распределяется между арендатором и землевладельцем.
В ситуации с наличными арендатор несет большую часть риска. В соглашении о разделе урожая производственный и ценовой риск распределяется между арендатором и землевладельцем.
Аренда за наличный расчет – один из наименее требовательных вариантов для собственника. Этот вид договоренностей включает в себя поиск арендатора, уплату налогов на недвижимость и несколько других мелких имущественных обязанностей.Ферма может использовать ставку арендной платы за всю ферму, которая будет включать всю землю на ферме. Или ферма может быть арендована за фиксированную сумму за акр пахотных земель с другой арендной ставкой для пастбищ или зданий. Как правило, арендные ставки на всю ферму ниже, чем арендная ставка на пахотные земли.
Определить справедливую арендную ставку непросто. Один из способов определения денежной арендной платы — установить ставку, аналогичную той, которую взимают другие люди в вашем регионе. Если вы используете этот метод, убедитесь, что вы сравниваете свои ставки с землевладельцами с аналогичным качеством земли. Всегда сравнивайте несколько ставок с одной или двумя. Сравнение арендных ставок имеет свои недостатки, поскольку предполагает, что другие взимают плату со своих арендаторов разумно и справедливо. Чтобы обеспечить справедливую цену, примите во внимание среднюю урожайность, рейтинг пригодности кукурузы (CSR2) и долю валовой стоимости урожая. В интересах всех участников поддерживать отношения между землевладельцем и арендатором в атмосфере партнерства, а не вражды.
Всегда сравнивайте несколько ставок с одной или двумя. Сравнение арендных ставок имеет свои недостатки, поскольку предполагает, что другие взимают плату со своих арендаторов разумно и справедливо. Чтобы обеспечить справедливую цену, примите во внимание среднюю урожайность, рейтинг пригодности кукурузы (CSR2) и долю валовой стоимости урожая. В интересах всех участников поддерживать отношения между землевладельцем и арендатором в атмосфере партнерства, а не вражды.
При долевой аренде арендатор и землевладелец делили часть расходов и урожая.Точное разделение является предметом переговоров. У землевладельцев также есть возможность гибкой аренды, которая может быть комбинацией как денежных средств, так и функций доли урожая.
Опять же, наилучшая договоренность об аренде зависит от каждой ситуации. В некоторых случаях может потребоваться рассмотрение других факторов, не связанных с арендованным имуществом, таких как уборка снега, уход за имуществом и техническое обслуживание.
Индивидуальное фермерство
Индивидуальное фермерство может служить альтернативой аренде фермы.Владелец земли принимает решения об урожае, ресурсах и маркетинге, но нанимает других для выполнения фактической работы. Таможенный оператор выполняет все операции с машиной на земле владельца в обмен на установленную плату или ставку. Индивидуальное земледелие — это вариант, сопряженный с большим риском для землевладельца. В плохой год прибыль от индивидуального земледелия будет меньше, чем при обычной аренде, но это компромисс, связанный с риском более высокой прибыли в некоторые годы.
Наем профессионального менеджера
Наем профессионального менеджера – это вариант для тех, кто хочет сохранить ферму в качестве инвестиции, но не хочет принимать деловые решения относительно фермы.Это расположение варьируется. В большинстве случаев владелец не принимает бизнес-решений и позволяет управлять фермой нанятому профессиональному управляющему фермой.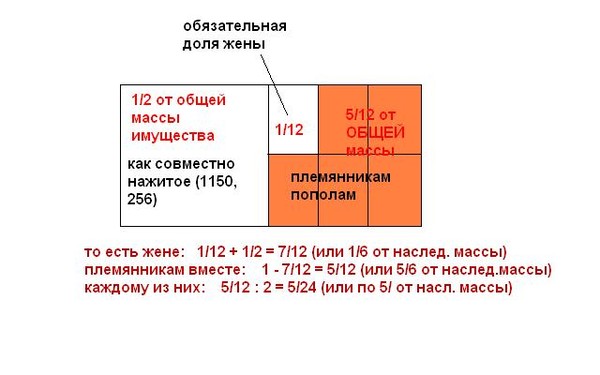 Владелец получает чек, а управляющая ферма получает процент от валового дохода. Типичная плата за денежную ренту составляет 3-6%, а доля урожая составляет около 10% валового дохода. Эти проценты будут варьироваться в зависимости от количества времени и усилий, затрачиваемых руководителем фермы.
Владелец получает чек, а управляющая ферма получает процент от валового дохода. Типичная плата за денежную ренту составляет 3-6%, а доля урожая составляет около 10% валового дохода. Эти проценты будут варьироваться в зависимости от количества времени и усилий, затрачиваемых руководителем фермы.
Возврат к земле как инвестиция обычно рассматривается в двух компонентах.Во-первых, это ежегодный денежный доход, который варьируется в зависимости от использования земли и типа договора аренды. Он подвержен погодным, производственным и ценовым рискам. В прошлом годовая денежная отдача от земли значительно варьировалась, в последние годы она составляла в среднем 2-3% после уплаты налога на имущество и до уплаты подоходного налога.
Вторым компонентом отдачи от земли является увеличение стоимости или прирост капитала. Средняя стоимость земли в США в 2020 году составляла 3160 долларов за акр. Это значение такое же, как и в 2019 году, но 5.на 3% выше, чем пять лет назад. Среднее значение измеряется стоимостью земли и построек на ферме. Стоимость земли сильно различается по стране и по типу сельскохозяйственных угодий. Стоимость пахотных земель в Соединенных Штатах в 2020 году оценивалась в 4100 долларов, а пастбища в среднем составляли 1400 долларов. Производительность и стоимость земли варьируются в зависимости от штата. С диапазоном стоимости государственных пахотных земель в 2020 году от 1030 до 13 800 долларов.
Стоимость земли сильно различается по стране и по типу сельскохозяйственных угодий. Стоимость пахотных земель в Соединенных Штатах в 2020 году оценивалась в 4100 долларов, а пастбища в среднем составляли 1400 долларов. Производительность и стоимость земли варьируются в зависимости от штата. С диапазоном стоимости государственных пахотных земель в 2020 году от 1030 до 13 800 долларов.
Независимо от используемого метода аренды настоятельно рекомендуется заключить письменный договор между землевладельцем и арендатором, чтобы каждая сторона имела четкое представление о соглашении.
Продажа земли
Последний вариант — продать землю. Этот вариант особенно привлекателен для тех, кто хочет использовать доходы от наследства для других целей. Это важно не только для сторон, но и их наследников.
При принятии решения о продаже недвижимости необходимо учитывать множество соображений. Налоговые последствия продажи зависят от нескольких факторов: суммы продажи, базы налога на прибыль имущества, а иногда и способа продажи. Продажа имущества по контракту может растянуть налогооблагаемую прибыль на несколько лет, что снижает налоговую нагрузку, но откладывает использование средств для других целей. В некоторых штатах действуют специальные программы по налоговым льготам для людей, продающих землю начинающему фермеру.
Продажа имущества по контракту может растянуть налогооблагаемую прибыль на несколько лет, что снижает налоговую нагрузку, но откладывает использование средств для других целей. В некоторых штатах действуют специальные программы по налоговым льготам для людей, продающих землю начинающему фермеру.
Те, кто решит продать землю, должны рассмотреть способ продажи. Большинство используют лицензированного брокера по недвижимости. Другие наследники продают землю тому, кто знаком с землей, или через рекламу.
Еще один способ продать ферму — провести земельный аукцион. Неясно, какой метод дает самую высокую цену; это зависит от участников торгов. Установление минимальной цены — один из вариантов для тех, кто проводит земельные аукционы. Если аукцион дает более высокую цену, чем заранее установленная минимальная цена, земля продается. Однако, если минимальная цена не достигнута, владелец может пересмотреть метод продажи или свои ожидания. Можно разделить большую собственность на более мелкие участки для аукциона. Например, наследники могут решить продать участок под застройку и оставить за собой пахотные земли, или наоборот.
Например, наследники могут решить продать участок под застройку и оставить за собой пахотные земли, или наоборот.
Многие наследники, решившие продать землю, не знают хорошей продажной цены. Рынок земли не имеет четкого определения по сравнению с другими более часто продаваемыми товарами. Наследники сельскохозяйственных угодий должны получить профессиональную оценку, чтобы определить стоимость земли и установить правильную цену продажи. Также важно помнить, что сентиментальная привязанность наследника к ферме не превращается в ценность для кого-то другого.Также важно не забывать обращаться за квалифицированной юридической консультацией.
Продажа за наличный расчет
Продажа за наличный расчет — один из самых популярных способов продажи земли. Продажа за наличные немедленно производит выручку от продажи для других целей. Это также сводит к минимуму риск для продавца. Однако продажа за наличные может сократить количество потенциальных покупателей, поскольку большинству покупателей потребуется какое-либо финансирование. Меньшее количество покупателей может означать более низкую цену продажи.
Меньшее количество покупателей может означать более низкую цену продажи.
Продажа по договору
Продажа по договору – это вариант, при котором покупатель вносит первоначальный взнос, а затем производит платежи за землю.Преимущество для покупателя заключается в том, что ему нужно меньше денег вперед. К преимуществам продавца относится возможность отчитываться о прибыли в рассрочку и снижать общие налоговые последствия. Договор купли-продажи подвергает продавца риску невыполнения обязательств покупателем. Однако повышенный риск означает, что продавец обычно может получить более высокие проценты. Более высокий процент является дополнительным доходом для продавца. Продавец может получить более высокую процентную ставку, чем банк заплатил бы за компакт-диск. С практической точки зрения большинство продавцов предпочли бы более высокую цену продажи (прирост капитала) и более низкую процентную ставку (обычный доход), а покупатель, конечно, хочет обратного. Опять же, в некоторых штатах есть специальные программы для начинающих фермеров, которые допускают различные налоговые режимы для этого дохода. Кроме того, предложение выгодного контракта увеличит число потенциальных покупателей и даст возможность получить более выгодную цену.
Опять же, в некоторых штатах есть специальные программы для начинающих фермеров, которые допускают различные налоговые режимы для этого дохода. Кроме того, предложение выгодного контракта увеличит число потенциальных покупателей и даст возможность получить более выгодную цену.
Обмен
Обмен недвижимостью — еще один вариант, который может быть привлекательным в некоторых обстоятельствах. Это особенно актуально для наследника, который уже занимается сельским хозяйством в другом районе. Продав имущество тому, кто владеет или может купить имущество ближе к существующей ферме, наследник может избежать налогов, которые могут быть получены от продажи.Обмен сельхозугодий на другую арендуемую недвижимость — еще одна возможность.
Подарок
Дарение имущества некоммерческой организации является еще одной альтернативой его продаже. Обычно это происходит, когда собственность имеет уникальную дикую природу или живописную ценность и желательна для таких организаций, как Nature Conservancy, Ducks Unlimited или Pheasants Forever. Этот вариант также позволяет наследникам избежать налоговых последствий.
Этот вариант также позволяет наследникам избежать налоговых последствий.
Как ферма была унаследована
В одиночку
Самостоятельное наследование фермы является более простым из двух сценариев.В этом случае любые решения будут основываться на обстоятельствах конкретного наследника без учета ожиданий и желаний других.
Наследование фермы в качестве физического лица по-прежнему оставляет те же основные варианты и альтернативы: продать ферму, сохранить ее в качестве инвестиции или обрабатывать землю.
Если наследник в настоящее время занимается сельским хозяйством, решение о том, что делать с унаследованной фермой, зависит от того, насколько ферма вписывается в текущую операцию и цели. В зависимости от того, как ферма была унаследована, ее можно консолидировать путем продажи унаследованной фермы и покупки фермы ближе к дому, не вызывая налоговых обязательств.Обсудить варианты с доверенным налоговым консультантом необходимо, прежде чем определять какие-либо преимущества использования Раздела 1031 Налогового кодекса.
Если наследник в настоящее время не занимается сельским хозяйством, применяются ранее обсужденные подводные камни и соображения. Наследники должны помнить, что не следует позволять романтическим представлениям о сельском хозяйстве или сентиментальной ценности земли затуманивать их мысли и оценивать ожидаемый доход из всех источников.
Если наследник оставляет землю в качестве инвестиции, он или она должны решить, сколько времени и энергии посвятить этому.В зависимости от того, как с этим обращаться, владение землей может потребовать значительно больше времени, чем обычные активы. Наследники сельскохозяйственных угодий также должны определить, сколько и какой риск они готовы нести.
Индивидуальные налоговые последствия, вероятно, будут определять решения о продаже земли. Существует множество альтернатив, и их следует тщательно рассмотреть. Кроме того, существуют варианты отсрочки или устранения налогового бремени в зависимости от того, как продается недвижимость.
С другими
Наследование фермы с другими людьми усложняет принятие решений.У каждого наследника разные цели и потребности. Однако слепое преследование того, что отвечает интересам одного наследника, может снизить прибыль для всех. Наследники должны думать как группа и стремиться к компромиссам. Открытое общение имеет важное значение.
В идеале человек, оставивший ферму наследникам, сделал все возможное для справедливости. Однако в некоторых случаях справедливое распределение являлось направляющей силой. В этих случаях справедливость становится индивидуальным соображением.
Осложнение возникает, если фермой в настоящее время управляет один из наследников.Важно помнить, что решения, принятые наследниками, не занимающимися сельским хозяйством, могут существенно повлиять на способность наследника, занимающегося сельским хозяйством, продолжать свою деятельность. Если наследники захотят продать, наследник-фермер может не иметь капитала или быть в состоянии взять на себя или купить всю ферму в одиночку. Это также может быть правдой, если один из наследников хочет заняться сельским хозяйством, а другие нет.
Это также может быть правдой, если один из наследников хочет заняться сельским хозяйством, а другие нет.
Если принято решение оставить ферму в качестве инвестиционной собственности, наследники должны договориться о том, как это сделать. Во многих случаях профессиональному управляющему фермой легче справиться с инвестициями, чтобы избежать осложнений.Независимо от выбранного метода должны быть установлены некоторые средства принятия решений. Один наследник не должен иметь возможности влиять на возврат инвестиций без согласия других.
Заключение
Наследование фермы может быть захватывающим и полезным, но в то же время это может быть значительный объем работы.
Точная профессиональная оценка определит размер наследства. ISU Extension and Outreach и другие организации могут помочь с решениями о вариантах и альтернативах для унаследованной фермы.
Наследники должны помнить, что могут быть существенные различия в налоговых последствиях различных вариантов. Перед принятием решения о том, что делать с унаследованной фермой, необходимо получить профессиональную налоговую и юридическую консультацию.
Перед принятием решения о том, что делать с унаследованной фермой, необходимо получить профессиональную налоговую и юридическую консультацию.
В конце концов, вы и другие, унаследовавшие землю, должны принять окончательное решение. Наиболее важными факторами, которые следует учитывать при принятии решения о том, что делать с фермой, являются ваши ценности и цели.
Ресурсы
ISU Extension and Outreach — Ag, принимающий решения
Землеустроительная экспертиза штата Айова
Центр сельскохозяйственного права и налогообложения
Почва и землепользование
Начальный фермерский центр
Обследование владения и владения сельскохозяйственными угодьями в штате Айова, 1982–2017 годы: перспектива за 35 лет
Национальная служба сельскохозяйственной статистики Министерства сельского хозяйства США
Сводка стоимости земли USDA NASS
Энн М.Йоханнс, специалист по программам распространения сельскохозяйственных знаний, 515-337-2766, aholste@iastate. edu
edu
Кельвин Лейболд, специалист по управлению фермерскими хозяйствами, 641-648-4850, [email protected]
Среднее наследство и пять советов, как оставить одно наследнику
Для многих получение наследства может показаться сбывшейся мечтой. С другой стороны, передача наследства может быть менее чем захватывающим опытом, если не делать это осторожно.
Исследование, проведенное Ameriprise Financial, показало, что, хотя 83 процента людей хотят оставить наследство, только 64 процента считают, что они на пути к этому.Возможно, это связано с масштабностью задачи. Существует не только необходимость оставить своим наследникам наследство выше среднего, но и неуверенность в том, что вы должны оставить разным людям.
Это руководство расскажет вам о среднем наследстве, подскажет, как оставить его и как узнать, когда настало подходящее время для передачи ваших активов.
Ключевые результаты:- Среднее наследство в США в период с 2016 по 2019 год составляло 46 200 долларов .
- Наследство от родителей с высшим образованием более чем на 10 000 долларов больше, чем наследство от родителей без образования.
- Только унаследованных поместий стоимостью более 11 700 000 долларов США облагаются налогом на наследство , но наследники могут нести ответственность за уплату налога на дарение в отношении всего, что превышает 15 000 долларов США.

Среднее наследство в США
Среднее наследство от родителей, бабушек и дедушек или других благотворителей в США составляет примерно 46 200 долларов, также согласно Обзору потребительских финансов.Среднее значение для одного процента самых богатых достигает 719 000 долларов, в то время как среднее значение для следующих девяти процентов резко снижается до 174 200 долларов.
- Средний размер наследства: 46 200 долларов
- Среднее ожидаемое наследство: 72 200 долларов
Помимо среднего наследования, данные Федеральной резервной системы также подчеркивают ключевые различия в среднем наследовании между расами и этническими группами:
- Медиана наследования среди белых семей: 88 500 долларов
- Медиана наследства среди чернокожих семей: 85 800 долларов США
- Медиана наследства среди латиноамериканских семей: 52 200 долларов США
Накопленные данные Организации экономического сотрудничества и развития показывают, что унаследованное богатство в США. С начала 1900-х годов на С. приходилось примерно от 50 до 60 процентов частного богатства.
С начала 1900-х годов на С. приходилось примерно от 50 до 60 процентов частного богатства.
Наследство от родителей
Согласно последним данным Федеральной резервной системы, средний размер наследства, полученного от родителей с высшим образованием, с 2016 по 2019 год составлял 92 700 долларов. Это на 16 500 долларов больше, чем среднее наследство, полученное от родителей без высшего образования.
Точно так же дети родителей со степенью также в два раза чаще ожидают наследства.Только 9,5% лиц, у которых родители не имеют высшего образования, рассчитывают на получение наследства, в то время как 23,6% лиц, у которых есть родители со степенью, рассчитывают получить имущество, переданное им по наследству.
Налоги с наследства
В зависимости от типа наследства требуемые налоги различаются. Например, в большинстве случаев наследники не обязаны подавать декларацию о налоге на наследство при наследовании имущества. Это верно, если только это имущество не превысит 11 700 000 долларов в 2021 году, по данным Налоговой службы.
Хотя наследники могут не облагаться налогом на наследство, вам, вероятно, придется заплатить налог на дарение. По данным IRS, с 2018 по 2021 год годовой предел освобождения от этого налога составляет 15 000 долларов.
Если вы не уверены, как будет выглядеть ваш налог на наследство, поговорите с финансовым консультантом или другим специалистом в области финансов, который может дать вам лучшее представление о том, чего ожидать.
Что считается большим наследством?
Существуют различные размеры наследства, но общее практическое правило: 100 000 долларов или более считаются крупным наследством.Получение такой значительной суммы денег потенциально может пугать, особенно если вам никогда раньше не приходилось распоряжаться такими деньгами.
Прежде чем потратить их на новую спортивную машину или загородный дом, подумайте, как извлечь максимальную пользу из своего наследства. Вы можете решить потратить их на эти предметы, но вам следует сесть с финансовым консультантом, чтобы обсудить ваши варианты и составить план, прежде чем делать какой-либо быстрый выбор.
Консультант может помочь вам понять, сколько вы должны инвестировать, откладывать на сбережения и свободно тратить.Они также могут посоветовать вам, является ли покупка аннуитета для гарантированного дохода в будущем хорошим вариантом для вас.
Пять советов, как оставить наследство своим близким
Оставить наследство своим наследникам — это процесс, который, скорее всего, не произойдет в одночасье. Найдите время, чтобы подумать, кто больше всего выиграет от определенных активов, и как лучше всего передать эти предметы каждому человеку.
Также следует подумать о том, сколько давать своим наследникам — будь то большое, малое или среднее наследство.Целесообразно также определить индивидуальные потребности и уровень понимания денег каждым человеком. Это может сильно повлиять на то, сколько вы жертвуете или как вы это делаете.
1. Управляйте своими ожиданиями
Прежде всего, вы должны понять, какие активы вы можете позволить себе оставить. Обязательно вычтите все пенсионные сбережения или другие финансовые средства, на которые вы будете полагаться до самой смерти.
Обязательно вычтите все пенсионные сбережения или другие финансовые средства, на которые вы будете полагаться до самой смерти.
Всегда лучше давать реалистично, чем давать чрезмерно щедро.Если вы в конечном итоге отдадите слишком много, ваши наследники могут использовать свои унаследованные деньги для оплаты ваших расходов в конце жизни. Это может привести к семейным спорам и напряженности вокруг того, кто должен нести ответственность за оплату этих счетов позже.
2. Оцените понимание наследником денег
Прежде чем подарить наследнику 1 миллион долларов или даже 50 000 долларов, вы должны убедиться, что он не потратит их без планирования. Чтобы лучше понять, как ваш наследник справится с крупной суммой денег, дайте ему небольшую пробную партию с несколькими тысячами долларов.Наблюдайте, инвестируют ли они их, тратят или выплачивают долги.
На что бы они ни использовали деньги, это даст вам хорошее представление о том, как они могут использовать большие суммы денег. Если они используют его с умом, вы можете сразу же подарить деньги. Однако, если они не совсем понимают, как эффективно использовать деньги, вы можете рассмотреть возможность размещения денег в доверительном управлении или покупки отсроченной ренты с единой премией на их имя.
Если они используют его с умом, вы можете сразу же подарить деньги. Однако, если они не совсем понимают, как эффективно использовать деньги, вы можете рассмотреть возможность размещения денег в доверительном управлении или покупки отсроченной ренты с единой премией на их имя.
Если пройти пробный запуск не представляется возможным, задайте себе следующие вопросы, чтобы лучше понять отношения ваших наследников с деньгами.
- Они тратят деньги на прихоти?
- Они полагаются на долг?
- У них есть стабильная работа?
- Они придерживаются бюджета каждый месяц?
- У них есть деньги, отложенные на непредвиденные расходы?
3. Рассмотрите варианты
Хотя вы можете дать своим наследникам единовременную сумму денег, есть и другие способы передать наследство. Как упоминалось ранее, не каждый метод подходит для каждого человека. Из-за этого вы должны знать все доступные вам варианты, в том числе:
- Рот ИРА
- Инвестиционный счет, где можно указать другое лицо в качестве бенефициара
- Ежегодные денежные взносы в виде подарков
- Каждый год делать денежные подарки, чтобы избежать высоких налогов на крупную сумму денег
Существует множество способов оставить любимому человеку наследство. Создание плана недвижимости поможет вам понять, что лучше для них, и может помочь вам определить метод передачи вашего имущества.
Создание плана недвижимости поможет вам понять, что лучше для них, и может помочь вам определить метод передачи вашего имущества.
4. Управляйте ожиданиями своих наследников
Легко увлечься, когда знаешь, что есть возможность унаследовать любой тип активов. Однако это может вызвать разочарование и разочарование, если вы заранее не установите четких ожиданий.
Подумайте, какие ожидания могут быть у ваших наследников, и используйте их, чтобы вести вдумчивый разговор с каждым из близких.Заблаговременная подготовка к проблемам, в том числе к потенциальному разочарованию в связи с наследством меньше среднего, может помочь вам узнать, как наилучшим образом справиться с ситуацией.
Имейте в виду, что вам не нужно явно раскрывать, что вы даете каждому человеку и сколько вы жертвуете на благотворительность или другие организации. Однако важно объяснить своим наследникам, на что они реально могут рассчитывать.
5. Общайтесь четко
Четкое общение может впоследствии устранить путаницу и враждебность между близкими. С кем вы решите поделиться своей финансовой информацией, в конечном итоге зависит от вас. Однако, если наследство ваших детей выглядит иначе, объяснение ваших рассуждений каждому ребенку в отдельности может иметь большое значение.
С кем вы решите поделиться своей финансовой информацией, в конечном итоге зависит от вас. Однако, если наследство ваших детей выглядит иначе, объяснение ваших рассуждений каждому ребенку в отдельности может иметь большое значение.
Когда давать деньги по наследству своим детям или внукам
Решение о передаче наследства зависит от множества факторов, но единого правильного ответа не существует. Каждая ситуация индивидуальна и к ней следует относиться соответственно. Если вы даете наследство нескольким людям, определите лучшее время для каждого человека, чтобы получить его.
Например, если вы планируете дать по 1 миллиону долларов каждому из ваших двух детей, рассмотрите их обстоятельства, характер и умение обращаться с деньгами в отдельности. Скажем, ваш старший ребенок изо всех сил пытается сохранить работу и всегда просит денег. Вместо того, чтобы давать их наследство сразу, подумайте о том, чтобы распределить его в течение определенного периода времени.
Точно так же, если ваш младший ребенок свободно и успешно обращается с деньгами, возможно, отдать его наследство единовременно было бы разумным решением.
Принимая решение о передаче денег в наследство своим детям или внукам, будьте осторожны и не делайте этого до того, как умрете. Ожидание освобождения их унаследованных активов после вашей смерти может помочь вам убедиться, что у вас достаточно ресурсов для оплаты ухода в конце жизни и связанных с этим расходов.
Независимо от того, оставляете ли вы наследство выше среднего, ниже среднего или среднее, найдите время, чтобы спланировать наилучший метод и время для его передачи. После этого вы можете решить пойти против своей первоначальной мысли и аннуитетировать деньги или передать их в доверительное управление.Что бы вы ни решили, убедитесь, что это правильное решение для вас и вашей семьи.
Генетика — Медицинская микробиология — Книжная полка NCBI
Генетическая информация у микробов
Генетический материал бактерий и плазмид — ДНК. Бактериальные вирусы
(бактериофаги или фаги) имеют ДНК или РНК в качестве генетического материала. Два основных
функциями генетического материала являются репликация и экспрессия. Генетический материал должен
точно воспроизвести, чтобы потомство наследовало все специфические генетические
детерминанты (генотип) родительского организма.Экспрессия специфических генетических
материал при определенном наборе условий роста определяет наблюдаемую
характеристики (фенотип) организма. Бактерии имеют мало структурных или
особенности развития, которые можно легко наблюдать, но они имеют широкий спектр
биохимические возможности и модели чувствительности к противомикробным агентам или
бактериофаги. Эти последние характеристики часто выбираются в качестве унаследованных
признаки, подлежащие анализу в исследованиях бактериальной генетики.
Два основных
функциями генетического материала являются репликация и экспрессия. Генетический материал должен
точно воспроизвести, чтобы потомство наследовало все специфические генетические
детерминанты (генотип) родительского организма.Экспрессия специфических генетических
материал при определенном наборе условий роста определяет наблюдаемую
характеристики (фенотип) организма. Бактерии имеют мало структурных или
особенности развития, которые можно легко наблюдать, но они имеют широкий спектр
биохимические возможности и модели чувствительности к противомикробным агентам или
бактериофаги. Эти последние характеристики часто выбираются в качестве унаследованных
признаки, подлежащие анализу в исследованиях бактериальной генетики.
Структура нуклеиновой кислоты
Нуклеиновые кислоты представляют собой крупные полимеры, состоящие из повторяющихся нуклеотидных единиц ().Каждый нуклеотид содержит один
фосфатная группа, один пентозный или дезоксипентозный сахар и один пуриновый или пиримидиновый
база. В ДНК сахар представляет собой D-2-дезоксирибозу; в РНК сахаром является D-рибоза. В ДНК
пуриновые основания – это аденин (А) и гуанин (G), а пиримидиновые основания – это
тимин (Т) и цитозин (С). В РНК урацил (U) заменяет тимин. Химически
модифицированные пуриновые и пиримидиновые основания обнаружены у некоторых бактерий и
бактериофаги. Повторяющаяся структура полинуклеотидов включает чередующиеся
остатки сахара и фосфата, с фосфодиэфирными связями, связывающими
3′-гидроксильная группа одного нуклеотидного сахара к 5′-гидроксильной
группа соседнего нуклеотидного сахара.Эти асимметричные фосфодиэфирные связи
определяют полярность полинуклеотидной цепи. пуриновое или пиримидиновое основание
связаны у 1′-атома углерода каждого остатка сахара и выступают из
повторяющийся сахарофосфатный остов. Двухцепочечная ДНК спиральна, и
две нити в спирали антипараллельны. Двойная спираль стабилизируется
водородные связи между пуриновыми и пиримидиновыми основаниями на противоположных цепях. В
каждой позиции, А на одной нити соединяется двумя водородными связями с Т на
противоположная цепь, или G соединяется тремя водородными связями с C.
В ДНК сахар представляет собой D-2-дезоксирибозу; в РНК сахаром является D-рибоза. В ДНК
пуриновые основания – это аденин (А) и гуанин (G), а пиримидиновые основания – это
тимин (Т) и цитозин (С). В РНК урацил (U) заменяет тимин. Химически
модифицированные пуриновые и пиримидиновые основания обнаружены у некоторых бактерий и
бактериофаги. Повторяющаяся структура полинуклеотидов включает чередующиеся
остатки сахара и фосфата, с фосфодиэфирными связями, связывающими
3′-гидроксильная группа одного нуклеотидного сахара к 5′-гидроксильной
группа соседнего нуклеотидного сахара.Эти асимметричные фосфодиэфирные связи
определяют полярность полинуклеотидной цепи. пуриновое или пиримидиновое основание
связаны у 1′-атома углерода каждого остатка сахара и выступают из
повторяющийся сахарофосфатный остов. Двухцепочечная ДНК спиральна, и
две нити в спирали антипараллельны. Двойная спираль стабилизируется
водородные связи между пуриновыми и пиримидиновыми основаниями на противоположных цепях. В
каждой позиции, А на одной нити соединяется двумя водородными связями с Т на
противоположная цепь, или G соединяется тремя водородными связями с C. Две нити
Следовательно, двухспиральные ДНК комплементарны. Из-за комплементарности,
двухцепочечная ДНК содержит эквимолярные количества пуринов (А + G) и
пиримидины (T + C), где A равно T, а G равно C, но моль
доля G + C в ДНК сильно различается у разных бактерий.
Информация в нуклеиновых кислотах закодирована упорядоченной последовательностью нуклеотидов.
по полинуклеотидной цепи, а в двухцепочечной ДНК последовательность каждого
цепь определяет, какой должна быть последовательность комплементарной цепи.То
степень гомологии последовательностей между ДНК разных микроорганизмов
самый строгий критерий для определения того, насколько тесно они связаны.
Две нити
Следовательно, двухспиральные ДНК комплементарны. Из-за комплементарности,
двухцепочечная ДНК содержит эквимолярные количества пуринов (А + G) и
пиримидины (T + C), где A равно T, а G равно C, но моль
доля G + C в ДНК сильно различается у разных бактерий.
Информация в нуклеиновых кислотах закодирована упорядоченной последовательностью нуклеотидов.
по полинуклеотидной цепи, а в двухцепочечной ДНК последовательность каждого
цепь определяет, какой должна быть последовательность комплементарной цепи.То
степень гомологии последовательностей между ДНК разных микроорганизмов
самый строгий критерий для определения того, насколько тесно они связаны.
Рисунок 5-1
Двойная спиральная структура ДНК. На схеме показана структура ДНК, представленная в виде спиральной лестница. Скелет каждой полинуклеотидной нити (представлен в виде лента) состоит из чередующихся остатков фосфата и дезоксирибозы связанные фосфодиэфиром (подробнее…)
Репликация ДНК
Во время репликации бактериального генома каждая нить двойной спирали ДНК
служит матрицей для синтеза новой комплементарной цепи. Каждая дочь
Таким образом, двухцепочечная молекула ДНК содержит одну старую полинуклеотидную цепь и одну
вновь синтезированная цепь. Такой тип репликации ДНК называется
полуконсервативный. Репликация хромосомной ДНК у бактерий начинается
определенный участок хромосомы, называемый началом, и продолжается в обоих направлениях до тех пор, пока
процесс завершен (). Когда
бактерии делятся бинарным делением после завершения репликации ДНК, т.
реплицированные хромосомы делятся на каждую из дочерних клеток. То
области происхождения специфически и временно связываются с клеточной мембраной
после того, как репликация ДНК была инициирована, что привело к модели, в которой мембрана
прикрепление направляет разделение дочерних хромосом (модель репликона).Эти характеристики репликации ДНК во время роста бактерий соответствуют
Требования к генетическому материалу должны быть точно воспроизведены и
наследуется каждой дочерней клеткой во время клеточного деления.
Каждая дочь
Таким образом, двухцепочечная молекула ДНК содержит одну старую полинуклеотидную цепь и одну
вновь синтезированная цепь. Такой тип репликации ДНК называется
полуконсервативный. Репликация хромосомной ДНК у бактерий начинается
определенный участок хромосомы, называемый началом, и продолжается в обоих направлениях до тех пор, пока
процесс завершен (). Когда
бактерии делятся бинарным делением после завершения репликации ДНК, т.
реплицированные хромосомы делятся на каждую из дочерних клеток. То
области происхождения специфически и временно связываются с клеточной мембраной
после того, как репликация ДНК была инициирована, что привело к модели, в которой мембрана
прикрепление направляет разделение дочерних хромосом (модель репликона).Эти характеристики репликации ДНК во время роста бактерий соответствуют
Требования к генетическому материалу должны быть точно воспроизведены и
наследуется каждой дочерней клеткой во время клеточного деления.
Рисунок 5-2
Авторадиограмма интактной реплицирующейся хромосомы E. палочка . Бактерии были радиоактивно мечены тритиевым тимидином для
примерно два поколения и были осторожно лизированы. Бактериальная ДНК
Затем исследовали авторадиографию. Вставка показывает репликацию
бактериальные (подробнее..)
палочка . Бактерии были радиоактивно мечены тритиевым тимидином для
примерно два поколения и были осторожно лизированы. Бактериальная ДНК
Затем исследовали авторадиографию. Вставка показывает репликацию
бактериальные (подробнее..)
Генная экспрессия
Генетическая информация, закодированная в ДНК, выражается путем синтеза специфических РНК и
белки, и информация течет от ДНК к РНК к белку. ДНК-ориентированный
синтез РНК называется транскрипцией. Поскольку нити двойной спирали
ДНК антипараллельны и комплементарны, только одна из двух цепей ДНК может
служить матрицей для синтеза определенной молекулы мРНК. Мессенджер РНК
(мРНК) передают информацию от ДНК, и каждая мРНК у бактерий функционирует как
матрица для синтеза одного или нескольких специфических белков.Процесс
нуклеотидная последовательность молекулы мРНК которой определяет первичную аминогруппу.
кислотная последовательность белка называется трансляцией. Рибосомы, комплексы
рибосомные РНК (рРНК) и несколько рибосомных белков переводят каждую мРНК в
соответствующую полипептидную последовательность с помощью транспортных РНК (тРНК),
аминоацил-тРНК-синтезазы, факторы инициации и факторы элонгации. Все
эти компоненты аппарата синтеза белка функционируют в
образование множества различных белков.Ген – это последовательность ДНК, которая кодирует
белок, рРНК или молекула тРНК (продукт гена).
Все
эти компоненты аппарата синтеза белка функционируют в
образование множества различных белков.Ген – это последовательность ДНК, которая кодирует
белок, рРНК или молекула тРНК (продукт гена).
Генетический код определяет, как нуклеотиды в мРНК определяют аминокислоты
в полипептиде. Потому что в мРНК всего 4 разных нуклеотида.
(содержащие U, A, C и G), одиночные нуклеотиды не содержат достаточной информации
однозначно указать все 20 аминокислот. В динуклеотидах 16 (4
× 4) расположение четырех нуклеотидов возможно, и в
возможно расположение тринуклеотидов 64 (4 × 4 × 4).Таким образом, требуется минимум три нуклеотида, чтобы обеспечить хотя бы один уникальный
последовательности, соответствующей каждой из 20 аминокислот. То
«Универсальный» генетический код, используемый большинством организмов (), представляет собой триплетный код, в котором 61 из
64 возможных тринуклеотида (кодона) кодируют определенные аминокислоты, и любой из
три оставшихся кодона (UAG, UAA или UGA) приводят к терминации
перевод. Кодоны, обрывающие цепь, также называют нонсенс-кодонами.
потому что в них не указаны аминокислоты. Генетический код описывается как
вырожденными, потому что несколько кодонов могут использоваться для одной аминокислоты, и как
неперекрывающиеся, так как соседние кодоны не имеют общих нуклеотидов.Исключения из «универсального» кода включают использование UGA в качестве
кодон триптофана у некоторых видов Mycoplasma и у
митохондриальная ДНК и несколько дополнительных различий кодонов в митохондриальной ДНК
из дрожжей, Drosophila и млекопитающих. Трансляция мРНК
обычно инициируется кодоном AUG для метионина, а соседние кодоны
транслируется последовательно по мере того, как мРНК считывается с 5′ по 3′
направление. Соответствующая полипептидная цепь собирается, начиная с
амино-концу и идет к его карбокси-концу.Последовательность амино
кислот в полипептиде, таким образом, совпадает с последовательностью
нуклеотидов в мРНК и соответствующий ген. Специфические ферментативные реакции
участвуют в синтезе ДНК, РНК и белка, выходят за рамки этого
глава.
Кодоны, обрывающие цепь, также называют нонсенс-кодонами.
потому что в них не указаны аминокислоты. Генетический код описывается как
вырожденными, потому что несколько кодонов могут использоваться для одной аминокислоты, и как
неперекрывающиеся, так как соседние кодоны не имеют общих нуклеотидов.Исключения из «универсального» кода включают использование UGA в качестве
кодон триптофана у некоторых видов Mycoplasma и у
митохондриальная ДНК и несколько дополнительных различий кодонов в митохондриальной ДНК
из дрожжей, Drosophila и млекопитающих. Трансляция мРНК
обычно инициируется кодоном AUG для метионина, а соседние кодоны
транслируется последовательно по мере того, как мРНК считывается с 5′ по 3′
направление. Соответствующая полипептидная цепь собирается, начиная с
амино-концу и идет к его карбокси-концу.Последовательность амино
кислот в полипептиде, таким образом, совпадает с последовательностью
нуклеотидов в мРНК и соответствующий ген. Специфические ферментативные реакции
участвуют в синтезе ДНК, РНК и белка, выходят за рамки этого
глава.
Экспрессия генетических детерминант у бактерий включает однонаправленный поток информации от ДНК к РНК к белку. У бактериофагов либо ДНК, либо РНК может служить генетическим материалом. При заражении бактерий РНК бактериофагов молекулы РНК служат матрицами для репликации РНК и мРНК.Исследования с группой ретровирусов вирусов животных показывают, что ДНК молекулы могут быть синтезированы из матриц РНК с помощью ферментов, обозначаемых как РНК-зависимые ДНК-полимеразы (обратные транскриптазы). Это обращение Обычное направление потока генетической информации от РНК к ДНК, а не от ДНК в РНК — важный механизм передачи информации от ретровирусов. кодироваться в ДНК и встраиваться в геномы животных клетки.
Организация генома
Молекулы ДНК, которые реплицируются как дискретные генетические единицы в бактериях, называются
репликоны.В некоторых штаммах Escherichia coli хромосома является
единственный репликон, присутствующий в клетке. Другие бактериальные штаммы имеют дополнительные
репликоны, такие как плазмиды и бактериофаги.
Хромосомная ДНК
Бактериальные геномы различаются по размеру примерно от 0,4 × 10 9 до 8,6
× 10 9 дальтонов (Да), некоторые из самых маленьких являются обязательными
паразиты ( Mycoplasma ) и самые крупные, принадлежащие к бактериям
способны к сложной дифференциации, такие как Myxococcus .То
количество ДНК в геноме определяет максимальное количество информации, которую он
может кодировать. Большинство бактерий имеют гаплоидный геном, состоящий из одной хромосомы.
кольцевой двухцепочечной молекулы ДНК. Однако линейные хромосомы имеют
были обнаружены у грамположительных боррелий и Streptomyces spp., а также один линейный и один круговой
хромосома присутствует у грамотрицательной бактерии Agrobacterium
тумефациенс . Единственная хромосома общего кишечника
бактерия Е.coli составляет 3 × 10 9 Да (4500
пар килобаз [kbp]) размером, что составляет от 2 до 3
процентов от сухой массы клетки. Геном E. coli всего около 0,1% размера генома человека, но этого достаточно для кодирования
несколько тысяч полипептидов среднего размера (40 кДа или 360 аминокислот).
Геном E. coli всего около 0,1% размера генома человека, но этого достаточно для кодирования
несколько тысяч полипептидов среднего размера (40 кДа или 360 аминокислот).
Хромосома E. coli имеет контурную длину приблизительно
1,35 мм, в несколько сотен раз длиннее бактериальной клетки, но ДНК
суперскручены и плотно упакованы в бактериальный нуклеоид.Требуемое время
для репликации всей хромосомы составляет около 40 минут, что составляет
примерно вдвое меньше кратчайшего времени деления этой бактерии. ДНК
репликация должна начинаться так же часто, как клетки делятся, поэтому быстро
растущих бактерий новый цикл хромосомной репликации начинается раньше предыдущего
раунд завершен. При быстром росте может быть четыре хромосомы.
репликация с образованием восьми во время клеточного деления, что связано с
завершение цикла хромосомной репликации.Таким образом, хромосома в
быстрорастущие бактерии размножаются более чем в одной точке. Репликация
Хромосомная ДНК у бактерий сложна и включает в себя множество различных
белки.
Плазмиды
Плазмиды представляют собой репликоны, которые сохраняются как дискретные внехромосомные генетические
элементы в бактериях. Обычно они намного меньше, чем бактериальные.
хромосомы, варьируя от менее 5 до более чем нескольких сотен т.п.н., хотя
плазмиды размером до 2 Мб встречаются у некоторых бактерий. Плазмиды обычно кодируют
признаки, которые не являются существенными для жизнеспособности бактерий, и реплицируются
независимо от хромосомы.Большинство плазмид имеют суперскрученную кольцевую форму.
молекулы двухцепочечной ДНК, но также были продемонстрированы линейные плазмиды.
в Borrelia и Streptomyces . Тесно связанные
или идентичные плазмиды демонстрируют несовместимость; они не могут быть стабильно
сохраняется в одном и том же бактериальном хозяине. Классификация плазмид основана на
несовместимости или при использовании специфических ДНК-зондов в тестах гибридизации для
определить последовательности нуклеотидов, характерные для конкретной плазмиды
репликоны. Некоторые гибридные плазмиды содержат более одного репликона. Конъюгативные
плазмиды кодируют функции, которые способствуют передаче плазмиды от донора
бактерия к другим бактериям-реципиентам, но неконъюгативные плазмиды — нет.
Конъюгативные плазмиды, которые также способствуют переносу бактериальной хромосомы из
донорную бактерию другим бактериям-реципиентам называют плазмидами фертильности,
и обсуждаются ниже. Среднее число молекул данной плазмиды на
бактериальной хромосомы называется ее числом копий.Крупные плазмиды (>40
пары килобаз) часто сопряжены, имеют небольшое число копий (от 1 до нескольких на
хромосомы), код для всех функций, необходимых для их репликации, и
распределяются между дочерними клетками во время клеточного деления таким образом,
похож на бактериальную хромосому. Плазмиды размером менее 7,5 тысяч пар оснований
обычно неконъюгативны, имеют большое число копий (обычно 10–20
на хромосому), полагаются на своего бактериального хозяина, чтобы обеспечить некоторые функции, необходимые
для репликации и случайным образом распределяются между дочерними клетками в
разделение.
Некоторые гибридные плазмиды содержат более одного репликона. Конъюгативные
плазмиды кодируют функции, которые способствуют передаче плазмиды от донора
бактерия к другим бактериям-реципиентам, но неконъюгативные плазмиды — нет.
Конъюгативные плазмиды, которые также способствуют переносу бактериальной хромосомы из
донорную бактерию другим бактериям-реципиентам называют плазмидами фертильности,
и обсуждаются ниже. Среднее число молекул данной плазмиды на
бактериальной хромосомы называется ее числом копий.Крупные плазмиды (>40
пары килобаз) часто сопряжены, имеют небольшое число копий (от 1 до нескольких на
хромосомы), код для всех функций, необходимых для их репликации, и
распределяются между дочерними клетками во время клеточного деления таким образом,
похож на бактериальную хромосому. Плазмиды размером менее 7,5 тысяч пар оснований
обычно неконъюгативны, имеют большое число копий (обычно 10–20
на хромосому), полагаются на своего бактериального хозяина, чтобы обеспечить некоторые функции, необходимые
для репликации и случайным образом распределяются между дочерними клетками в
разделение.
Многие плазмиды контролируют важные с медицинской точки зрения свойства патогенных бактерий, включая резистентность к одному или нескольким антибиотикам, выработку токсинов и синтез структур клеточной поверхности, необходимых для прикрепления или колонизации. Плазмиды, определяющие устойчивость к антибиотикам, часто называют R-плазмидами. (или R-факторы). Типичные токсины, кодируемые плазмидами, включают термолабильные и термостабильные энтеротоксины E. coli , эксфолиативный токсин Staphylococcus aureus и столбнячный токсин Столбнячная клостридия .Некоторые плазмиды скрыты и не имеют заметное воздействие на бактериальные клетки, в которых они обитают. Сравнение плазмиды профили — это полезный метод для оценки возможного родства отдельных клинические изоляты определенного вида бактерий для эпидемиологических исследований. Обсуждается роль плазмид в эволюции устойчивости к антибиотикам. ниже.
Бактериофаги
Бактериофаги (бактериальные вирусы, фаги) – инфекционные агенты, способные размножаться
как облигатные внутриклеточные паразиты у бактерий. Внеклеточные фаговые частицы
метаболически инертны и состоят в основном из белков и нуклеиновых кислот
(ДНК или РНК, но не то и другое одновременно). Белки фаговой частицы образуют защитную
оболочка (капсид), окружающая плотно упакованный геном нуклеиновой кислоты. Фаг
геномы различаются по размеру примерно от 2 до 200 килобаз на нить нуклеиновой кислоты.
кислоты и состоят из двухцепочечной ДНК, одноцепочечной ДНК или РНК. Фаг
геномы, как и плазмиды, кодируют функции, необходимые для репликации у бактерий.
но в отличие от плазмид они также кодируют капсидные белки и неструктурные белки.
требуется для сборки фага.Несколько морфологически различных типов фагов
описаны, в том числе полиэдрические, нитевидные и сложные. Сложный
фаги имеют многогранные головки, к которым присоединяются хвосты, а иногда и другие придатки (хвост
пластинки, хвостовые волокна и др.) прикрепляются.
Внеклеточные фаговые частицы
метаболически инертны и состоят в основном из белков и нуклеиновых кислот
(ДНК или РНК, но не то и другое одновременно). Белки фаговой частицы образуют защитную
оболочка (капсид), окружающая плотно упакованный геном нуклеиновой кислоты. Фаг
геномы различаются по размеру примерно от 2 до 200 килобаз на нить нуклеиновой кислоты.
кислоты и состоят из двухцепочечной ДНК, одноцепочечной ДНК или РНК. Фаг
геномы, как и плазмиды, кодируют функции, необходимые для репликации у бактерий.
но в отличие от плазмид они также кодируют капсидные белки и неструктурные белки.
требуется для сборки фага.Несколько морфологически различных типов фагов
описаны, в том числе полиэдрические, нитевидные и сложные. Сложный
фаги имеют многогранные головки, к которым присоединяются хвосты, а иногда и другие придатки (хвост
пластинки, хвостовые волокна и др.) прикрепляются.
Один цикл роста фага показан на . Инфекция инициируется адсорбцией фага на специфических
рецепторы на поверхности восприимчивых бактерий-хозяев. Капсиды остаются на
клеточной поверхности, и геномы ДНК или РНК проникают в клетки-мишени (проникновение).Поскольку инфекционность геномной ДНК или РНК намного меньше, чем у зрелых
вируса, есть время сразу после заражения, называемое периодом затмения
во время которого внутриклеточный инфекционный фаг не может быть обнаружен. Заражение
фаговая РНК или ДНК реплицируются для производства множества новых копий фагового генома,
и продуцируются фаг-специфические белки. Для большинства фагов сборка потомства
происходит в цитоплазме, а высвобождение потомства происходит путем лизиса клеток. В
напротив, нитчатые фаги образуются в клеточной оболочке и высвобождаются
не убивая клетки-хозяева.Период затмения заканчивается, когда внутриклеточно
появляется инфекционное потомство. Латентный период – это время от заражения
до появления внеклеточного потомства, а период нарастания – это интервал от
конец латентного периода, пока все фаги не станут внеклеточными. Среднее число
фаговых частиц, продуцируемых каждой инфицированной клеткой, называется размером вспышки.
Капсиды остаются на
клеточной поверхности, и геномы ДНК или РНК проникают в клетки-мишени (проникновение).Поскольку инфекционность геномной ДНК или РНК намного меньше, чем у зрелых
вируса, есть время сразу после заражения, называемое периодом затмения
во время которого внутриклеточный инфекционный фаг не может быть обнаружен. Заражение
фаговая РНК или ДНК реплицируются для производства множества новых копий фагового генома,
и продуцируются фаг-специфические белки. Для большинства фагов сборка потомства
происходит в цитоплазме, а высвобождение потомства происходит путем лизиса клеток. В
напротив, нитчатые фаги образуются в клеточной оболочке и высвобождаются
не убивая клетки-хозяева.Период затмения заканчивается, когда внутриклеточно
появляется инфекционное потомство. Латентный период – это время от заражения
до появления внеклеточного потомства, а период нарастания – это интервал от
конец латентного периода, пока все фаги не станут внеклеточными. Среднее число
фаговых частиц, продуцируемых каждой инфицированной клеткой, называется размером вспышки. характерен для каждого вируса и часто колеблется от 50 до нескольких сотен.
Для обсуждения структуры, размножения и классификации животных
вирусы, см. главы 41 и 42.
характерен для каждого вируса и часто колеблется от 50 до нескольких сотен.
Для обсуждения структуры, размножения и классификации животных
вирусы, см. главы 41 и 42.
Рисунок 5-3
Одностадийный рост бактериофага. Культуру восприимчивых бактерий синхронно заражают бактериофаг добавляют в момент времени 0 при низкой множественности заражения. Вскоре после этого неабсорбированный фаг инактивируют добавлением антифаговая антисыворотка, (подробнее…)
Фаги подразделяются на две основные группы: вирулентные и умеренные. Рост
вирулентные фаги в восприимчивых бактериях разрушают клетки-хозяева. Заражение
чувствительные бактерии фагами умеренного пояса могут иметь один из двух исходов: литический
роста или лизогении.Литический рост умеренных и вирулентных бактериофагов
сходны, что приводит к образованию фагового потомства и гибели бактерий-хозяев.
Лизогения — это особый тип латентной вирусной инфекции, при котором фаговый геном
размножается как профаг в бактериальной клетке. У большинства лизогенных бактерий
гены, необходимые для развития литических фагов, не экспрессируются, и продукция
инфекционный фаг не встречается. Кроме того, лизогенные клетки невосприимчивы к
суперинфекция вирусом, который они укрывают в виде профага.Физическое состояние
профага не одинакова для всех умеренных вирусов. Например,
интегрирован профаг бактериофага λ в E. coli в бактериальную хромосому в определенном месте и реплицируется как часть
бактериальной хромосоме, тогда как профаг бактериофага Р1 в E.
coli реплицируется как внехромосомная плазмида.
У большинства лизогенных бактерий
гены, необходимые для развития литических фагов, не экспрессируются, и продукция
инфекционный фаг не встречается. Кроме того, лизогенные клетки невосприимчивы к
суперинфекция вирусом, который они укрывают в виде профага.Физическое состояние
профага не одинакова для всех умеренных вирусов. Например,
интегрирован профаг бактериофага λ в E. coli в бактериальную хромосому в определенном месте и реплицируется как часть
бактериальной хромосоме, тогда как профаг бактериофага Р1 в E.
coli реплицируется как внехромосомная плазмида.
Рост литического фага происходит спонтанно в небольшой части лизогенных клеток,
и несколько внеклеточных фагов присутствуют в культурах лизогенных бактерий.Для некоторых лизогенных бактерий синхронная индукция развития литических фагов
происходит во всей популяции лизогенных бактерий при обработке их
агенты, повреждающие ДНК, такие как ультрафиолетовый свет или митомицин С. Потеря
профаг из лизогенной бактерии, переводя ее в нелизогенное состояние и
восстановление восприимчивости к заражению фагом, который изначально присутствовал
как профаг, называется лечением.
Некоторые фаги умеренного пояса содержат гены бактериальных характеристик, которые
не связанный с развитием литического фага или лизогенным состоянием, и экспрессией
такие гены называют фаговой конверсией (или лизогенной конверсией).Примеры
конверсия фагов, которая важна для микробной вирулентности, включает продукцию
токсина дифтерии Corynebacterium diphtheriae ,
эритрогенный токсин Streptococcus pyogenes (группа А
β-гемолитические стрептококки), ботулинический токсин Clostridium
botulinum и шига-подобные токсины E. coli . В
В каждом из этих примеров ген, кодирующий бактериальный токсин, присутствует в
геном умеренного фага. Специфичность О-антигенов в Salmonella также можно контролировать с помощью фаговой конверсии.Фаготипирование – это тестирование штаммов определенного вида бактерий на
чувствительность к специфическим бактериофагам. Модели восприимчивости к
набор типирующих фагов предоставляет информацию о возможном родстве
отдельные клинические изоляты. Такая информация особенно полезна для
эпидемиологические расследования.
Такая информация особенно полезна для
эпидемиологические расследования.
Мутация и селекция
Мутации — это наследуемые изменения в геноме. Спонтанные мутации у отдельных
бактерии встречаются редко. Некоторые мутации вызывают изменения фенотипических характеристик; в
о возникновении таких мутаций можно судить по эффектам, которые они вызывают.В
специфические для микробной генетики эталонные организмы обозначаются как штаммы дикого типа,
а потомки, имеющие мутации в геноме, называются мутантами. Таким образом,
мутанты характеризуются унаследованными различиями между ними и их
предковые штаммы дикого типа. Вариантными формами определенной генетической детерминанты являются
называются аллелями. Генотипические символы — это строчные буквы, выделенные курсивом аббревиатуры.
укажите отдельные гены с надстрочным индексом (+), указывающим на дикий тип
аллель. Фенотипические символы написаны с заглавной буквы, а не курсивом, чтобы различать их.
из генотипических символов.Например, генотипический символ способности производить
β-галактозидаза, необходимая для ферментации лактозы. lacZ + и мутанты, которые не могут производить
β-галактозидаза lacZ . Фенотип ферментации лактозы
обозначается Lac + , а неспособность ферментировать лактозу —
Лак – .
lacZ + и мутанты, которые не могут производить
β-галактозидаза lacZ . Фенотип ферментации лактозы
обозначается Lac + , а неспособность ферментировать лактозу —
Лак – .
Обнаружение мутантных фенотипов
Селективные и дифференциальные среды полезны для выделения бактериальных мутантов.
Некоторые селективные среды позволяют расти определенным мутантам, но не позволяют
штаммы дикого типа для роста.Редкие мутанты могут быть выделены с помощью таких селективных
средства массовой информации. Дифференциальные среды позволяют расти и формироваться бактериям дикого типа и мутантам.
колонии, различающиеся по внешнему виду. Обнаружение редких мутантов на дифференциалах
media ограничивается общим количеством наблюдаемых колоний. Учитывать
штамм дикого типа E. coli , чувствительный к
антибиотик стрептомицин (фенотип Str s ) и может использовать лактозу в качестве
единственный источник углерода (фенотип Lac + ). Спонтанно
встречающиеся мутанты Str r встречаются редко и обычно встречаются с частотой
менее одной на 109 бактерий в культурах дикого типа E.
палочка . Тем не менее, мутанты Strr могут быть легко выделены с помощью
селективная среда, содержащая стрептомицин, поскольку Str s дикого типа
бактерии погибают. Выделение лактозонегативных (фенотип
Lac – ) мутанты E. coli представляют собой
другая проблема. На минимальных средах с лактозой в качестве единственного источника углерода
Lac + штаммы дикого типа будут расти, но
Мутанты Lac – не могут расти.На дифференциальных средах, таких как
Агар МакКонки-лактоза или агар эозин-метиленовый синий-лактоза, Lac+
дикого типа и Lac – мутантных штаммов E.
coli можно отличить по цвету, но спонтанно
Мутанты Lac – слишком редки, чтобы их можно было легко выделить.
Селективная среда для Lac – мутантов E.
coli можно получить, включив химические аналоги лактозы, которые
превращаются в токсичные метаболиты бактериями Lac + , но
не мутантами Lac – .
Спонтанно
встречающиеся мутанты Str r встречаются редко и обычно встречаются с частотой
менее одной на 109 бактерий в культурах дикого типа E.
палочка . Тем не менее, мутанты Strr могут быть легко выделены с помощью
селективная среда, содержащая стрептомицин, поскольку Str s дикого типа
бактерии погибают. Выделение лактозонегативных (фенотип
Lac – ) мутанты E. coli представляют собой
другая проблема. На минимальных средах с лактозой в качестве единственного источника углерода
Lac + штаммы дикого типа будут расти, но
Мутанты Lac – не могут расти.На дифференциальных средах, таких как
Агар МакКонки-лактоза или агар эозин-метиленовый синий-лактоза, Lac+
дикого типа и Lac – мутантных штаммов E.
coli можно отличить по цвету, но спонтанно
Мутанты Lac – слишком редки, чтобы их можно было легко выделить.
Селективная среда для Lac – мутантов E.
coli можно получить, включив химические аналоги лактозы, которые
превращаются в токсичные метаболиты бактериями Lac + , но
не мутантами Lac – . Мутанты Lac — затем могут расти на таких средах, но бактерии дикого типа Lac + убиты.
Мутанты Lac — затем могут расти на таких средах, но бактерии дикого типа Lac + убиты.
Мутации, инактивирующие основные гены у гаплоидных организмов, обычно летальные, но такие потенциально летальные мутации часто можно изучать, если их выражение контролируется путем манипулирования условиями эксперимента. За например, мутация, повышающая термолабильность эссенциального гена продукт может предотвращать рост бактерий при 42°C, хотя мутантный Бактерии все еще могут расти при 25°C.И наоборот, чувствительные к холоду мутанты экспрессируют мутантный фенотип при низкой температуре, но не при высокой температуре. Мутации, чувствительные к температуре и холоду, являются примерами условных мутации, как и подавляемые мутации, описанные далее в этой главе. А условно-летальный фенотип указывает на то, что мутантный ген необходим для жизнеспособность.
Спонтанные и индуцированные мутации
Частота мутаций у бактерий определяется точностью репликации ДНК,
возникновение повреждений ДНК и эффективность механизмов восстановления
поврежденной ДНК.
Для конкретного бактериального штамма при определенных условиях роста мутация скорость для любого конкретного гена постоянна и выражается как вероятность мутация за клеточное деление. В популяции бактерий, выращенных из небольшого инокулята доля мутантов обычно прогрессивно увеличивается по мере увеличения размера популяция бактерий увеличивается.
Мутации у бактерий могут возникать спонтанно и независимо от
экспериментальные методы, применяемые для их обнаружения. Этот принцип был впервые продемонстрирован
флуктуационным тестом ().То
количество устойчивых к фагам мутантов E. coli в повторах
культуры, выращенные из небольшого инокулята, измеряли и сравнивали с таковыми в
несколько образцов, взятых из одной культуры. Если мутации устойчивости к фагам
возникало только после воздействия фага, вариабельность числа мутантов
между культурами должны быть одинаковыми в обоих наборах условий. В отличие,
если устойчивые к фагам мутации возникали спонтанно до воздействия
бактерий в фаг, количество мутантов должно быть более изменчивым в
независимо выращенных культур, поскольку различия в размерах бактериальных
популяции, когда появился первый мутант, будет способствовать наблюдаемому
изменчивость. Данные показали, что мутации устойчивости к фагам в E. coli возник спонтанно с постоянной вероятностью
на клеточное деление.
Данные показали, что мутации устойчивости к фагам в E. coli возник спонтанно с постоянной вероятностью
на клеточное деление.
Рисунок 5-4
Испытание на колебания. Различия в количестве колоний фагорезистентных мутантов в повторные образцы из одной субкультуры были небольшими и отражали только ожидаемые колебания из-за ошибок выборки. В отличие, число устойчивых к фагам колоний (подробнее…)
Посев реплик подтвердил, что мутации в бактериях могут возникать спонтанно,
без воздействия на бактерии селективных агентов ().Для покрытия реплик, плоская, стерильная, вельветовая
поверхность используется для сбора инокулята с поверхности эталонной чашки с агаром
и перенесите образцы на другие чашки с агаром. Таким образом, образцы
бактериальная популяция с основной чашки переносится на чашки-реплики
не нарушая их пространственного расположения. Если пластины-реплики содержат
селективной среды и мастер-планшетов нет, положения выделенных мутантов
можно отметить колонии на чашках-репликах и бактерии, которые не подвергались воздействию
к селективным условиям можно выделить из одних и тех же позиций на
мастер пластина. Мутанты E. coli , устойчивые к бактериофагу Т1
или к стрептомицину были выделены таким образом, не подвергая
бактерии дикого типа к бактериофагу или антибиотику.
Мутанты E. coli , устойчивые к бактериофагу Т1
или к стрептомицину были выделены таким образом, не подвергая
бактерии дикого типа к бактериофагу или антибиотику.
Рисунок 5-5
Обнаружение ранее существовавших бактериальных мутантов по реплике покрытие. Мастер-планшет обильно инокулировали образцами чистых культур фагочувствительные бактерии. После инкубации бактерии от хозяина пластины были перенесены путем посева реплик на дубликаты (подробнее…)
На скорость мутаций влияют как экологические, так и генетические факторы.Контакт с
бактерий к мутагенным агентам вызывает увеличение частоты мутаций, иногда на
несколько порядков. Многие химические и физические агенты, в том числе рентгеновские лучи
и ультрафиолетового света, обладают мутагенной активностью. Химические вещества, которые являются канцерогенными
для животных часто являются мутагенными для бактерий или могут быть преобразованы животными
тканей до метаболитов, мутагенных для бактерий. Стандартизированные тесты для
мутагенность в бактериях используются в качестве процедур скрининга для выявления
агенты окружающей среды, которые могут быть канцерогенными для человека. Гены-мутаторы в
бактерии вызывают увеличение скорости спонтанных мутаций для широкого спектра
другие гены. Экспрессия этих генов, вызванная повреждением ДНК (см. ответ SOS
позже) позволяет восстанавливать повреждения ДНК, которые в противном случае были бы смертельными, но
подверженный ошибкам механизм, который увеличивает скорость мутации. Общий
частота мутаций — вероятность того, что мутация произойдет где-то в
бактериальный геном в расчете на клеточное деление — относительно постоянен для
разнообразие организмов с геномами разного размера и, по-видимому,
важный фактор в определении приспособленности бактериального штамма к выживанию
в природе.Большинство мутаций вредны, и риск неблагоприятных мутаций для
отдельные бактерии должны быть уравновешены положительным значением изменчивости, поскольку
механизм приспособления бактериальных популяций к изменяющимся условиям окружающей среды.
условия.
Гены-мутаторы в
бактерии вызывают увеличение скорости спонтанных мутаций для широкого спектра
другие гены. Экспрессия этих генов, вызванная повреждением ДНК (см. ответ SOS
позже) позволяет восстанавливать повреждения ДНК, которые в противном случае были бы смертельными, но
подверженный ошибкам механизм, который увеличивает скорость мутации. Общий
частота мутаций — вероятность того, что мутация произойдет где-то в
бактериальный геном в расчете на клеточное деление — относительно постоянен для
разнообразие организмов с геномами разного размера и, по-видимому,
важный фактор в определении приспособленности бактериального штамма к выживанию
в природе.Большинство мутаций вредны, и риск неблагоприятных мутаций для
отдельные бактерии должны быть уравновешены положительным значением изменчивости, поскольку
механизм приспособления бактериальных популяций к изменяющимся условиям окружающей среды.
условия.
Молекулярная основа мутаций
Мутации классифицируются на основе структурных изменений, происходящих в ДНК
(). Некоторые мутации являются
локализованы в пределах коротких сегментов ДНК (например, нуклеотидные замены,
микроделеции и микроинсерции).Другие мутации затрагивают большие области
ДНК и включают делеции, вставки или перестройки сегментов ДНК.
Некоторые мутации являются
локализованы в пределах коротких сегментов ДНК (например, нуклеотидные замены,
микроделеции и микроинсерции).Другие мутации затрагивают большие области
ДНК и включают делеции, вставки или перестройки сегментов ДНК.
Когда замена нуклеотида происходит в области ДНК, которая кодирует
полипептид, один из трех нуклеотидов в пределах одного кодона
соответствующая молекула мРНК будет изменена. Молчаливые мутации не вызывают изменений в
структуру или функцию полипептида, потому что один кодон в мРНК заменен на
другой для той же аминокислоты. Другие замены приводят к тому, что одна аминокислота становится
заменяется другим в определенном положении внутри полипептида
соответствующий измененному кодону.Мутации, приводящие к замещению одного
аминокислоты вместо другой в полипептидной цепи называются миссенс-мутациями.
Влияние замены аминокислот на функцию полипептидного гена
продукт варьируется и зависит от местоположения и идентичности аминокислоты
замена. Мутантные полипептиды, содержащие замены аминокислот, обычно
имеют общие антигенные детерминанты с полипептидом дикого типа и часто имеют некоторые
остаточной биологической активности. Мутации, приводящие к замене амино
кислотный кодон с терминирующим кодоном называют нонсенс-мутациями.Это приводит к
в продукции амино-концевого фрагмента нормального полипептида, когда
транслируется мутантная мРНК. Бессмысленные мутации часто приводят к полной потере
активность продукта гена.
Мутации, приводящие к замене амино
кислотный кодон с терминирующим кодоном называют нонсенс-мутациями.Это приводит к
в продукции амино-концевого фрагмента нормального полипептида, когда
транслируется мутантная мРНК. Бессмысленные мутации часто приводят к полной потере
активность продукта гена.
Из-за триплетной природы генетического кода последствия мутаций
вызваны вставками или делециями небольшого количества нуклеотидов
(микровставки, микроделеции) зависят как от количества, так и от последовательности
вовлечены нуклеотиды. Удаление или добавление кратных трем нуклеотидов
пары не влияет на рамку считывания, но вызывает удаление или добавление
соответствующее количество аминокислот в одном месте внутри полипептида.Если новый
кодон, обрывающий цепь, происходит преждевременное обрыв цепи
полипептид. Напротив, добавление или удаление других номеров
пары нуклеотидов изменяет рамку считывания для всего сегмента мРНК от
мутация дистального конца гена. Таким образом, мутации со сдвигом рамки считывания
может вызвать резкие изменения в структуре и активности полипептида. продукты генов, и их часто классифицируют как нонсенс-мутации.
продукты генов, и их часто классифицируют как нонсенс-мутации.
Тесты на комплементацию
Чтобы определить, локализованы ли мутации в одном и том же гене или в разных генах,
комплементарные тесты проводятся с частично диплоидными бактериальными штаммами
().Две копии области
бактериальной хромосомы с мутацией присутствуют в одном и том же
бактерия, причем каждая копия содержит другую мутацию (мутации находятся в
транс-аранжировка). Фенотип дикого типа указывает на то, что мутации находятся в
разные гены. Это явление называется комплементарностью. Если мутант
наблюдается фенотип, следует провести контрольный опыт с
мутации в цис-положении, чтобы исключить возможность того, что дикий тип
аллели не могут нормально экспрессироваться в частично диплоидном бактериальном штамме.Тесты комплемента первоначально назывались «цис-транс».
тесты, а термин цистрон иногда используется как синоним гена.
Тесты комплемента можно проводить и интерпретировать, даже если
биохимические функции продуктов гена неизвестны.
Рисунок 5-6
Комплементация — это метод проверки функционального гена продукты. Два мутанта со схожими фенотипами (неспособность конвертировать субстрат X к продукту Z). Мутации в этих штаммах обозначены а и б, соответственно, и аллели дикого типа (более…)
В качестве примера рассмотрим использование теста комплементарности для характеристики двух
независимо полученные мутанты Lac – E.
палочка . Биохимический путь утилизации лактозы требует
β-галактозид пермеаза (генотипический символ lac Y) для транспорта
лактозу в бактериальную клетку и β-галактозидазу (генотипический символ lacZ ) для превращения лактозы в D-глюкозу и D-галактозу. Мутанты
у которых отсутствует β-галактозидпермеаза или β-галактозидаза, не могут
использовать лактозу для роста.Если мутации в обоих Lac – мутанты инактивировали тот же белок (например, β-галактозид), затем
частично диплоидный штамм, содержащий гена lacZ обоих
мутанты в транс-положении не смогут утилизировать лактозу. В
напротив, если бы генотипы двух мутантов были lacZ + lacY и lacZ lacY + , частично диплоидная бактерия
будет продуцировать активную β-галактозидазу из lacZ + детерминантный и активный
β-галактозид проникает из lacY + определитель.Дополнение
произойдет, и частично диплоидный штамм будет использовать лактозу.
В
напротив, если бы генотипы двух мутантов были lacZ + lacY и lacZ lacY + , частично диплоидная бактерия
будет продуцировать активную β-галактозидазу из lacZ + детерминантный и активный
β-галактозид проникает из lacY + определитель.Дополнение
произойдет, и частично диплоидный штамм будет использовать лактозу.
Реверсия и подавление
Мутации, преобразующие фенотип из дикого типа в мутантный, называются прямыми
мутации и мутации, которые изменяют фенотип с мутантного обратно на дикий тип
называются обратными мутациями (реверсиями). Бактериальные штаммы, содержащие
обратные мутации называются ревертантами. Анализ мутаций, вызывающих
фенотипическая реверсия дает полезную информацию. Обратные мутации, которые восстанавливают
точная последовательность нуклеотидов ДНК дикого типа является истинной реверсией.Истинный
ревертанты идентичны штаммам дикого типа как генотипически, так и
фенотипически. Обратные мутации, которые не восстанавливают точный нуклеотид
последовательности ДНК дикого типа называются супрессорными мутациями (супрессорами).
Некоторые ревертанты, содержащие супрессорные мутации, фенотипически
неотличимы от штаммов дикого типа. Другие ревертанты, называемые
псевдоревертанты, можно фенотипически отличить от штаммов дикого типа,
например, тонкими различиями в характеристиках ферментативного
активность, которая была восстановлена (например, специфическая активность, субстрат
специфичность, кинетические константы или чувствительность к тепловым или химическим
инактивация).Распознавание псевдоревертантных фенотипов предполагает наличие
супрессорных мутаций.
Обратные мутации, которые не восстанавливают точный нуклеотид
последовательности ДНК дикого типа называются супрессорными мутациями (супрессорами).
Некоторые ревертанты, содержащие супрессорные мутации, фенотипически
неотличимы от штаммов дикого типа. Другие ревертанты, называемые
псевдоревертанты, можно фенотипически отличить от штаммов дикого типа,
например, тонкими различиями в характеристиках ферментативного
активность, которая была восстановлена (например, специфическая активность, субстрат
специфичность, кинетические константы или чувствительность к тепловым или химическим
инактивация).Распознавание псевдоревертантных фенотипов предполагает наличие
супрессорных мутаций.
Супрессорные мутации могут быть внутригенными или экстрагенными. Внутригенные супрессоры
расположены в том же гене, что и прямые мутации, которые они подавляют. То
возможные локализации и характер внутригенных супрессоров определяются
исходной прямой мутацией и отношениями между первичной структурой
продукта гена и его биологической активности. Экстрагенные супрессоры
расположенные в разных генах от мутаций, эффекты которых они подавляют.То
способность экстрагенных супрессоров подавлять множество независимых мутаций
можно протестировать. Некоторые экстрагенные супрессоры специфичны для определенных генов,
некоторые специфичны для определенных кодонов, а некоторые имеют другую специфичность
узоры. Экстрагенные супрессоры, обращающие фенотипические эффекты
Кодоны, обрывающие цепь, были хорошо охарактеризованы, и было обнаружено, что они изменяют
структура специфических тРНК. Конкретная супрессорная тРНК может позволить
кодон, обрывающий цепь, который транслируется, что приводит к включению
конкретной аминокислоты в формирующийся полипептид в положении, соответствующем
к кодону, обрывающему цепь.У бактерии с обрывом цепи
мутация и соответствующий экстрагенный супрессор, трансляция мРНК
содержащие мутантный кодон, могут привести к образованию полноразмерного
полипептид. Биологическая активность полноразмерного полипептида, образованного в виде
Последствия подавления зависят как от количества произведенного белка, так и от
функциональные последствия замены конкретной аминокислоты, определяемые
супрессорная тРНК.
Экстрагенные супрессоры
расположенные в разных генах от мутаций, эффекты которых они подавляют.То
способность экстрагенных супрессоров подавлять множество независимых мутаций
можно протестировать. Некоторые экстрагенные супрессоры специфичны для определенных генов,
некоторые специфичны для определенных кодонов, а некоторые имеют другую специфичность
узоры. Экстрагенные супрессоры, обращающие фенотипические эффекты
Кодоны, обрывающие цепь, были хорошо охарактеризованы, и было обнаружено, что они изменяют
структура специфических тРНК. Конкретная супрессорная тРНК может позволить
кодон, обрывающий цепь, который транслируется, что приводит к включению
конкретной аминокислоты в формирующийся полипептид в положении, соответствующем
к кодону, обрывающему цепь.У бактерии с обрывом цепи
мутация и соответствующий экстрагенный супрессор, трансляция мРНК
содержащие мутантный кодон, могут привести к образованию полноразмерного
полипептид. Биологическая активность полноразмерного полипептида, образованного в виде
Последствия подавления зависят как от количества произведенного белка, так и от
функциональные последствия замены конкретной аминокислоты, определяемые
супрессорная тРНК.
Обмен генетической информацией
Биологическое значение сексуальности у микроорганизмов заключается в увеличении вероятность того, что редкие, независимые мутации будут происходить вместе в одном микробе и подвергнуться естественному отбору.Генетические взаимодействия между микробами позволяют их геномы эволюционировали гораздо быстрее, чем путем одной мутации. Представитель явления, имеющие медицинское значение, которые связаны с обменом генетической информацией или Геномные перестройки включают быстрое появление и распространение антибиотиков. плазмиды резистентности, вариации жгутиковой фазы у сальмонелл и антигенные изменение поверхностных антигенов у Neisseria и Borrelia.
Половые процессы у бактерий включают передачу генетической информации от донора к
реципиента и приводят либо к замене аллелей донора на аллели реципиента
или добавление донорских генетических элементов к геному реципиента.трансформация,
трансдукция и конъюгация — это половые процессы, в которых используются разные механизмы для
ввести ДНК донора в бактерии-реципиенты (). Поскольку донорская ДНК не может сохраняться в реципиентной бактерии, если
это часть репликона, рекомбинация между геномами донора и реципиента часто
требуется для получения стабильного гибридного потомства. Рекомбинация наиболее вероятна
когда бактерии-доноры и реципиенты происходят из одного и того же или близкородственного
разновидность.
Поскольку донорская ДНК не может сохраняться в реципиентной бактерии, если
это часть репликона, рекомбинация между геномами донора и реципиента часто
требуется для получения стабильного гибридного потомства. Рекомбинация наиболее вероятна
когда бактерии-доноры и реципиенты происходят из одного и того же или близкородственного
разновидность.
Рисунок 5-7
Обмен генетической информацией у бактерий.Трансформация, трансдукция и конъюгация различаются по средствам для введение ДНК из клетки-донора в клетку-реципиент. А) В трансформации берутся фрагменты ДНК, высвобождаемые из бактерий-доноров компетентным (далее…)
Для обнаружения рекомбинанта его фенотип должен отличаться от обоих родительских
фенотипы. Для рекомбинантного фенотипа может потребоваться рост или деление клеток.
выражается. Задержка в выражении рекомбинантного фенотипа до гаплоидного
расщепление рекомбинантного генома называется задержкой сегрегации, и задержка до
синтез продуктов, кодируемых донорскими генами, называется фенотипическим отставанием. Тестирование на сцепление (неслучайная рекомбинация родительских аллелей в рекомбинантных
потомство) возможен, когда родительские бактерии имеют разные аллели для нескольких
гены. Донорский аллель невыбранного гена с большей вероятностью присутствует в
рекомбинантным, если он связан с выбранным донорным геном, чем если он не связан с
выбранного донорского гена. Количественный анализ сцепления позволяет построить
генетические карты. Геном E. coli является кольцевым (), что определяется как генетическими
сцепление и прямой биохимический анализ хромосомной ДНК, и генетическая карта
совпадает с физической картой хромосомной ДНК.Генетическое и физическое картирование
также используются для анализа внехромосомных репликонов, таких как бактериофаги и
плазмиды.
Тестирование на сцепление (неслучайная рекомбинация родительских аллелей в рекомбинантных
потомство) возможен, когда родительские бактерии имеют разные аллели для нескольких
гены. Донорский аллель невыбранного гена с большей вероятностью присутствует в
рекомбинантным, если он связан с выбранным донорным геном, чем если он не связан с
выбранного донорского гена. Количественный анализ сцепления позволяет построить
генетические карты. Геном E. coli является кольцевым (), что определяется как генетическими
сцепление и прямой биохимический анализ хромосомной ДНК, и генетическая карта
совпадает с физической картой хромосомной ДНК.Генетическое и физическое картирование
также используются для анализа внехромосомных репликонов, таких как бактериофаги и
плазмиды.
Рисунок 5-8
Круговая генетическая карта E. coli . Положения репрезентативных генов указаны во внутреннем круге.
Расстояния между генами откалиброваны в минутах на основе времени
требуется для передачи во время конъюгации.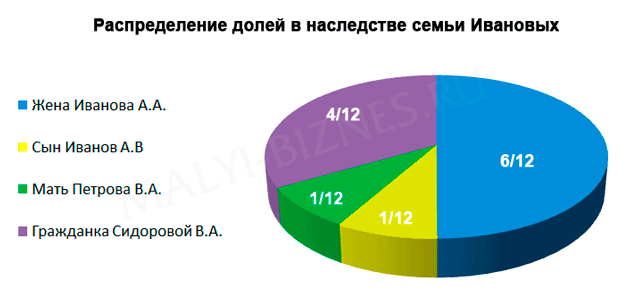 Позиция треонина (thr)
локус произвольно (подробнее…)
Позиция треонина (thr)
локус произвольно (подробнее…)
Многие бактерии имеют системы рестрикции-модификации (РМ), состоящие из модификации
ферменты, которые метилируют остатки аденина или цитозина в определенных последовательностях в их
собственной ДНК и соответствующих эндонуклеаз рестрикции, расщепляющих чужеродную ДНК, которая
не несет специфической модификации одних и тех же последовательностей-мишеней.Немного
Рестриктазы расщепляют только те ДНК, которые были метилированы в определенных местах.
последовательности. Эти рестрикционные системы, которые, возможно, эволюционировали для защиты бактерий
против инвазии фагами или плазмидами, являются важным барьером для генетических
обмены между различными бактериальными штаммами или видами. Недавние данные свидетельствуют
что переносимые плазмидами системы RM могут быть для плазмиды средством обеспечения ее переноса в
штамма-хозяина, поскольку клетки, потерявшие плазмиду (и соответствующий защитный
ген метилазы) погибают под действием более стабильного фермента рестрикции,
который атакует недавно реплицированную, но немодифицированную хромосомную ДНК.
Трансформация
При трансформации фрагменты ДНК, высвобождаемые из бактерий-доноров, поглощаются
непосредственно из внеклеточной среды бактериями-реципиентами. Рекомбинация
происходит между отдельными молекулами трансформирующей ДНК и хромосомами
реципиентные бактерии. Чтобы быть активными в трансформации, молекулы ДНК должны находиться на
не менее 500 нуклеотидов в длину, и трансформирующая активность быстро разрушается
путем обработки ДНК дезоксирибонуклеазой. Молекулы трансформирующей ДНК соответствуют
к очень маленьким фрагментам бактериальной хромосомы.Котрансформация генов
поэтому маловероятно, если только они не настолько тесно связаны, что их можно
кодируется одним фрагментом ДНК. Трансформация была обнаружена в Streptococcus pneumoniae и встречается в других бактериальных
роды, включая Haemophilus , Neisseria , Bacillus и Staphylococcus . Способность
Бактерии поглощают внеклеточную ДНК и трансформируются, что называется
компетентность, зависит от физиологического состояния бактерий. Многие бактерии
тех, кто обычно некомпетентен, можно заставить взять ДНК в лаборатории
манипуляции, такие как кальциевый шок или воздействие высоковольтного электрического
импульс (электропорация). У некоторых бактерий (включая Haemophilus и Neisseria)
Поглощение ДНК зависит от наличия специфических последовательностей олигонуклеотидов в
преобразование ДНК, но у других (включая Streptococcus pneumoniae) поглощение ДНК
не зависит от последовательности. Компетентные бактерии также могут поглощать интактные
ДНК бактериофага (трансфекция) или плазмидная ДНК, которая затем может реплицироваться как
внехромосомные генетические элементы в бактериях-реципиентах.Напротив,
часть хромосомной ДНК донорской бактерии обычно не может реплицироваться в
бактерия-реципиент, если только она не становится частью репликона в результате рекомбинации.
Исторически характеристика «преобразующего принципа»
из S. pneumoniae предоставил первое прямое доказательство того, что ДНК
генетический материал.
Многие бактерии
тех, кто обычно некомпетентен, можно заставить взять ДНК в лаборатории
манипуляции, такие как кальциевый шок или воздействие высоковольтного электрического
импульс (электропорация). У некоторых бактерий (включая Haemophilus и Neisseria)
Поглощение ДНК зависит от наличия специфических последовательностей олигонуклеотидов в
преобразование ДНК, но у других (включая Streptococcus pneumoniae) поглощение ДНК
не зависит от последовательности. Компетентные бактерии также могут поглощать интактные
ДНК бактериофага (трансфекция) или плазмидная ДНК, которая затем может реплицироваться как
внехромосомные генетические элементы в бактериях-реципиентах.Напротив,
часть хромосомной ДНК донорской бактерии обычно не может реплицироваться в
бактерия-реципиент, если только она не становится частью репликона в результате рекомбинации.
Исторически характеристика «преобразующего принципа»
из S. pneumoniae предоставил первое прямое доказательство того, что ДНК
генетический материал.
Трансдукция
При трансдукции бактериофаги действуют как векторы для введения ДНК от донора
бактерии в бактерии-реципиенты путем инфекции. Для некоторых фагов, называемых
генерализованные трансдуцирующие фаги, небольшая часть вирионов, продуцируемых во время
литический рост являются аберрантными и содержат случайный фрагмент бактериального генома
вместо фаговой ДНК.Каждый отдельный трансдуцирующий фаг несет различный набор
тесно связанных генов, представляющих небольшой сегмент генома бактерий.
Трансдукция, опосредованная популяциями таких фагов, называется генерализованной.
трансдукции, так как каждая часть бактериального генома имеет примерно
такая же вероятность передачи от донора к бактериям-реципиентам. Когда
генерализованный трансдуцирующий фаг заражает реципиентную клетку, экспрессия
происходит перенос донорских генов. Абортивная трансдукция относится к временному
экспрессия одного или нескольких донорских генов без образования рекомбинантного потомства,
тогда как полная трансдукция характеризуется производством стабильных
рекомбинанты, которые наследуют донорские гены и сохраняют способность к их экспрессии.В
абортивной трансдукции фрагмент ДНК донора не реплицируется, а среди
потомство исходного трансдуктана только одна бактерия содержит донорскую ДНК
фрагмент.
Для некоторых фагов, называемых
генерализованные трансдуцирующие фаги, небольшая часть вирионов, продуцируемых во время
литический рост являются аберрантными и содержат случайный фрагмент бактериального генома
вместо фаговой ДНК.Каждый отдельный трансдуцирующий фаг несет различный набор
тесно связанных генов, представляющих небольшой сегмент генома бактерий.
Трансдукция, опосредованная популяциями таких фагов, называется генерализованной.
трансдукции, так как каждая часть бактериального генома имеет примерно
такая же вероятность передачи от донора к бактериям-реципиентам. Когда
генерализованный трансдуцирующий фаг заражает реципиентную клетку, экспрессия
происходит перенос донорских генов. Абортивная трансдукция относится к временному
экспрессия одного или нескольких донорских генов без образования рекомбинантного потомства,
тогда как полная трансдукция характеризуется производством стабильных
рекомбинанты, которые наследуют донорские гены и сохраняют способность к их экспрессии.В
абортивной трансдукции фрагмент ДНК донора не реплицируется, а среди
потомство исходного трансдуктана только одна бактерия содержит донорскую ДНК
фрагмент. Во всех остальных потомках продукты донорских генов постепенно становятся
разводят после каждого поколения бактериального роста до тех пор, пока донорский фенотип не сможет
больше не выражаться. На селективной среде, на которой присутствуют только бактерии с
донорский фенотип может расти, абортивные трансдуктанты образуют мельчайшие колонии, которые
легко отличить от колоний стабильных трансдуктантов.Частота
абортивной трансдукции обычно на один-два порядка больше
чем частота генерализованной трансдукции, что указывает на то, что большинство клеток
инфицированные генерализованными трансдуцирующими фагами не продуцируют рекомбинантных
потомство.
Во всех остальных потомках продукты донорских генов постепенно становятся
разводят после каждого поколения бактериального роста до тех пор, пока донорский фенотип не сможет
больше не выражаться. На селективной среде, на которой присутствуют только бактерии с
донорский фенотип может расти, абортивные трансдуктанты образуют мельчайшие колонии, которые
легко отличить от колоний стабильных трансдуктантов.Частота
абортивной трансдукции обычно на один-два порядка больше
чем частота генерализованной трансдукции, что указывает на то, что большинство клеток
инфицированные генерализованными трансдуцирующими фагами не продуцируют рекомбинантных
потомство.
Специализированная трансдукция отличается от генерализованной трансдукции по нескольким параметрам.
Он опосредуется только специфическими фагами умеренного пояса и лишь несколькими специфическими донорами.
гены могут быть переданы бактериям-реципиентам. Специализированные трансдуцирующие фаги
образуются только тогда, когда лизогенные донорские бактерии входят в литический цикл и выделяют
фаговое потомство. Специализированные трансдуцирующие фаги представляют собой редкие рекомбинанты, которые
лишены части нормального фагового генома и содержат часть бактериального
хромосома, расположенная рядом с местом прикрепления профага. Многие специализированные
трансдуцирующие фаги дефектны и не могут завершить литический цикл фага
рост в инфицированных клетках, если только фаги-помощники не присутствуют, чтобы обеспечить недостающие
фаговые функции. Специализированная трансдукция возникает в результате лизогенизации
бактерия-реципиент специализированным трансдуцирующим фагом и экспрессией
донорские гены.Фаговая конверсия и специализированная трансдукция имеют много
сходство, но происхождение конвертирующих генов в умеренной конверсии
фагов неизвестно.
Специализированные трансдуцирующие фаги представляют собой редкие рекомбинанты, которые
лишены части нормального фагового генома и содержат часть бактериального
хромосома, расположенная рядом с местом прикрепления профага. Многие специализированные
трансдуцирующие фаги дефектны и не могут завершить литический цикл фага
рост в инфицированных клетках, если только фаги-помощники не присутствуют, чтобы обеспечить недостающие
фаговые функции. Специализированная трансдукция возникает в результате лизогенизации
бактерия-реципиент специализированным трансдуцирующим фагом и экспрессией
донорские гены.Фаговая конверсия и специализированная трансдукция имеют много
сходство, но происхождение конвертирующих генов в умеренной конверсии
фагов неизвестно.
Конъюгация
При конъюгации прямой контакт между бактериями-донорами и реципиентами приводит к
установление цитоплазматического мостика между ними и перенос части или всей
генома донора реципиенту. Способность донора определяется специфическими
конъюгативные плазмиды, называемые плазмидами фертильности или половыми плазмидами.
Плазмида F (также называемая фактором F) штамма E.coli является прототипом
для плазмид фертильности у грамотрицательных бактерий. Штаммы E.
coli с внехромосомной плазмидой F называются
F + и функционируют как доноры, тогда как штаммы, лишенные
Плазмиды F представляют собой F — и ведут себя как реципиенты. Спряжение
функции плазмиды F задаются кластером не менее чем из 25 трансферов
( tra ) гены, определяющие экспрессию F pili, синтез
и перенос ДНК во время спаривания, вмешательство в способность
Бактерии F + служат реципиентами и выполняют другие функции.Каждая бактерия F + имеет от 1 до 3 F-пилей, которые связываются со специфическими
белок внешней мембраны (продукт гена ompA) на бактериях-реципиентах для инициации
спаривание. Образуется межклеточный цитоплазматический мостик, и одна нить F
плазмидная ДНК передается от донора к реципиенту, начиная с уникального источника
и прогрессирует в направлении от 5′ к 3′. Переданный
цепь превращается в кольцевую двухцепочечную ДНК плазмиды F в реципиенте
бактерия, и в доноре синтезируется новая цепь, замещающая
переданная прядь.Обе эксконъюгирующие бактерии представляют собой F + ,
поэтому плазмида F может распространяться путем инфекции среди генетически совместимых
популяции бактерий. Помимо роли F-пилей в конъюгации,
они также функционируют как рецепторы для донор-специфических (мужских) фагов.
Переданный
цепь превращается в кольцевую двухцепочечную ДНК плазмиды F в реципиенте
бактерия, и в доноре синтезируется новая цепь, замещающая
переданная прядь.Обе эксконъюгирующие бактерии представляют собой F + ,
поэтому плазмида F может распространяться путем инфекции среди генетически совместимых
популяции бактерий. Помимо роли F-пилей в конъюгации,
они также функционируют как рецепторы для донор-специфических (мужских) фагов.
Плазмида F в E. coli может существовать как внехромосомная
генетический элемент или быть интегрированным в бактериальную хромосому (). Поскольку плазмида F и
бактериальная хромосома представляет собой обе кольцевые молекулы ДНК, реципрокная рекомбинация
между ними образуется больший ДНК-кольцо, состоящий из вставленной F-плазмидной ДНК.
линейно в хромосому. E. coli содержит несколько копий
нескольких различных генетических элементов, называемых инсерционными последовательностями (см.
транспозоны для более подробной информации), в различных местах его хромосомы и в
F плазмида. Гомологическая рекомбинация между последовательностями вставки в
хромосома и плазмида F приводит к предпочтительной интеграции плазмиды F
в хромосомных сайтах, где расположены инсерционные последовательности. хромосомный
Однако сайты, в которых обнаружены последовательности вставок, различаются среди штаммов Э.палочка .
Гомологическая рекомбинация между последовательностями вставки в
хромосома и плазмида F приводит к предпочтительной интеграции плазмиды F
в хромосомных сайтах, где расположены инсерционные последовательности. хромосомный
Однако сайты, в которых обнаружены последовательности вставок, различаются среди штаммов Э.палочка .
Рисунок 5-9
Роль плазмиды F в определении состояний донора и реципиента Кишечная палочка . Плазмида F является представителем специфических конъюгативных плазмид, которые контроль донорской способности в E. coli . F- штаммы отсутствуют плазмиду F и являются генетическими реципиентами. Питаются штаммы F+ (подробнее…)
Штамм E. coli с интегрированной плазмидой F сохраняет свой
способность функционировать в качестве донора в супружеских отношениях. Поскольку донор напрягается с
интегрированные F-факторы могут передавать хромосомные гены реципиентам с высоким
эффективности они называются Hfr ( H igh f requency r комбинация) штаммов. Перенос одноцепочечной ДНК
от донора Hfr к реципиенту начинается с начала внутри плазмиды F и
происходит, как описано выше, за исключением того, что перенесенная ДНК является гибридной
репликон, состоящий из плазмиды F, встроенной в бактериальную хромосому.
Перенос всего этого репликона, включая бактериальную хромосому, требует
примерно 100 минут. Идентичность первого хромосомного гена
переносится и полярность хромосомного переноса определяются сайтом
интеграции плазмиды F и ее ориентации по отношению к
бактериальная хромосома.Поскольку спаривающиеся бактерии обычно разделяются спонтанно.
перед переносом всей хромосомы конъюгация обычно передает
только фрагмент донорской хромосомы в реципиента. Вероятность того, что
ген донора попадет в бактерию-реципиент во время уменьшения конъюгации,
следовательно, как его расстояние от начала F (и, следовательно, время его
передача) увеличивается. Спаривающиеся клетки также могут быть разделены экспериментально с помощью
подвергая их сильному сдвиговому усилию в механическом смесителе; это
называется прерванным спариванием.
Перенос одноцепочечной ДНК
от донора Hfr к реципиенту начинается с начала внутри плазмиды F и
происходит, как описано выше, за исключением того, что перенесенная ДНК является гибридной
репликон, состоящий из плазмиды F, встроенной в бактериальную хромосому.
Перенос всего этого репликона, включая бактериальную хромосому, требует
примерно 100 минут. Идентичность первого хромосомного гена
переносится и полярность хромосомного переноса определяются сайтом
интеграции плазмиды F и ее ориентации по отношению к
бактериальная хромосома.Поскольку спаривающиеся бактерии обычно разделяются спонтанно.
перед переносом всей хромосомы конъюгация обычно передает
только фрагмент донорской хромосомы в реципиента. Вероятность того, что
ген донора попадет в бактерию-реципиент во время уменьшения конъюгации,
следовательно, как его расстояние от начала F (и, следовательно, время его
передача) увеличивается. Спаривающиеся клетки также могут быть разделены экспериментально с помощью
подвергая их сильному сдвиговому усилию в механическом смесителе; это
называется прерванным спариванием. Для образования рекомбинантного потомства требуется
рекомбинация между перенесенной ДНК донора и геномом реципиента
бактерия. Анализ потомства от спариваний, прерванных по разным причинам.
интервалы показывают, какие хромосомные гены передаются первыми
конкретных донорских штаммов, последовательное время проникновения генов, которые
передаваемые впоследствии, и все более низкая вероятность того, что гены
переданные позже, появятся в рекомбинантном потомстве. Округлость
генетическая карта E.coli первоначально был выведен из
перекрывающиеся, циклически переставленные группы связанных генов, которые были перенесены
рано отдельными донорскими штаммами, в которых фактор F был интегрирован в
различные хромосомные локации.
Для образования рекомбинантного потомства требуется
рекомбинация между перенесенной ДНК донора и геномом реципиента
бактерия. Анализ потомства от спариваний, прерванных по разным причинам.
интервалы показывают, какие хромосомные гены передаются первыми
конкретных донорских штаммов, последовательное время проникновения генов, которые
передаваемые впоследствии, и все более низкая вероятность того, что гены
переданные позже, появятся в рекомбинантном потомстве. Округлость
генетическая карта E.coli первоначально был выведен из
перекрывающиеся, циклически переставленные группы связанных генов, которые были перенесены
рано отдельными донорскими штаммами, в которых фактор F был интегрирован в
различные хромосомные локации.
При скрещивании F + и F – бактерий,
реципиентам с высокой эффективностью переносится только плазмида F.
Хромосомные гены передаются с очень низкой эффективностью, и это
спонтанные мутации Hfr в популяциях F + , которые опосредуют
перенос донорских хромосомных генов. В вязках между Hfr и
F – штаммы , сегмент плазмиды F, содержащий tra область переносится последней, после всего бактериального
хромосома была перенесена. Большинство рекомбинантов от скрещиваний между Hfr и
F — клетки не могут наследовать весь набор плазмиды F.
гены и фенотипически имеют F – . В вязках между
штаммы F + и F – , плазмида F
быстро распространяется в бактериальной популяции, и большинство рекомбинантов
Ф + .
В вязках между Hfr и
F – штаммы , сегмент плазмиды F, содержащий tra область переносится последней, после всего бактериального
хромосома была перенесена. Большинство рекомбинантов от скрещиваний между Hfr и
F — клетки не могут наследовать весь набор плазмиды F.
гены и фенотипически имеют F – . В вязках между
штаммы F + и F – , плазмида F
быстро распространяется в бактериальной популяции, и большинство рекомбинантов
Ф + .
Интегрированные плазмиды F в штаммах Hfr иногда могут быть вырезаны из бактериального
хромосома. Если вырезание точно обращает процесс интеграции, F+
вырабатываются клетки. Однако в редких случаях иссечение происходит путем
рекомбинации, включающие инсерционные последовательности или другие гены на бактериальном
хромосомы, расположенные на некотором расстоянии от исходного места интеграции.
В таких случаях сегменты бактериальной хромосомы могут включаться в
гибридные F-плазмиды, называемые F’-плазмидами (см. Ресурсы).Подобными процессами сегменты бактериального
Хромосома иногда может включаться в R-плазмиды для получения гибридных
R’ плазмиды. Конъюгативные плазмиды R’ могут функционировать как плазмиды фертильности, потому что
они могут интегрироваться в бактериальную хромосому путем гомологичной рекомбинации и
опосредует перенос хромосомных генов при спаривании с бактериями-реципиентами. Ф’
плазмиды, R’-плазмиды, специализированные трансдуцирующие фаги и рекомбинантные плазмиды
или фаги, сконструированные путем клонирования генов (описано ниже), представляют собой гибридные репликоны
которые могут включать сегменты бактериальной хромосомы.Следовательно, любой из этих
генетические элементы могут быть использованы для конструирования частично диплоидных бактериальных
штаммы, необходимые для комплементарных тестов и других целей.
Ресурсы).Подобными процессами сегменты бактериального
Хромосома иногда может включаться в R-плазмиды для получения гибридных
R’ плазмиды. Конъюгативные плазмиды R’ могут функционировать как плазмиды фертильности, потому что
они могут интегрироваться в бактериальную хромосому путем гомологичной рекомбинации и
опосредует перенос хромосомных генов при спаривании с бактериями-реципиентами. Ф’
плазмиды, R’-плазмиды, специализированные трансдуцирующие фаги и рекомбинантные плазмиды
или фаги, сконструированные путем клонирования генов (описано ниже), представляют собой гибридные репликоны
которые могут включать сегменты бактериальной хромосомы.Следовательно, любой из этих
генетические элементы могут быть использованы для конструирования частично диплоидных бактериальных
штаммы, необходимые для комплементарных тестов и других целей.
Конъюгация также происходит у грамположительных бактерий. Грамположительные донорские бактерии
продуцируют адгезины, которые заставляют их агрегировать с клетками-реципиентами, но половые пили
не участвуют. У некоторых видов Streptococcus реципиент
Бактерии продуцируют внеклеточные половые феромоны, вызывающие изменение фенотипа донора.
экспрессироваться бактериями, содержащими соответствующую конъюгативную плазмиду, и
конъюгативная плазмида не позволяет донорским клеткам производить соответствующий
феромон.
Рекомбинация
Рекомбинация включает разрыв и соединение родительских молекул ДНК с образованием гибридные, рекомбинантные молекулы. Было выявлено несколько различных видов рекомбинации. идентифицированы, которые зависят от различных особенностей участвующих геномов и требуют активности различных генных продуктов. Специфические ферменты, действующие на ДНК (например, экзонуклеазы, эндонуклеазы, полимеразы, лигазы) участвуют в рекомбинации. Подробное обсуждение биохимических событий рекомбинации выходит за рамки данной главы.
Генерализованная рекомбинация включает молекулы ДНК донора и реципиента, которые
гомологичные последовательности нуклеотидов. Взаимный обмен может происходить между любыми
гомологичные сайты донора и реципиента. В E. coli продукт
гена recA необходим для генерализованной рекомбинации,
но другие генные продукты также участвуют.
В E. coli продукт
гена recA необходим для генерализованной рекомбинации,
но другие генные продукты также участвуют.
Местная рекомбинация включает взаимные обмены только между специфическими
сайтов в молекулах ДНК донора и реципиента. Ген recA продукт не требуется для сайт-специфической рекомбинации.Интеграция
умеренный бактериофаг l в хромосому E. coli представляет собой
хорошо изученный пример сайт-специфической рекомбинации (). Специфические сайты прикрепления (att) на хромосома E. coli и ДНК фага λ имеют общие
коровая последовательность из 15 нуклеотидов, внутри которой происходит реципрокная рекомбинация,
фланкированы соседними последовательностями, которые не гомологичны в фаге и бактерии
геномы. У фага λ продукт гена int (интеграза) требуется для события интеграции для конкретного сайта в
лизогенизация; продукты int и xis (эксцизионных) генов необходимы для
комплементарное специфическое для сайта событие иссечения, которое происходит во время индукции литического
Развитие фагов в лизогенных клетках.
Рисунок 5-10
Интеграция и удаление бактериофага λ примеры сайт-специфической рекомбинации. λ ДНК показана тонкими линиями, а хромосомная ДНК – толстыми. линии. Сайты прикрепления (att) представляют собой закрытые ящики для бактерий. хромосомы и открытые ящики (подробнее…)
Незаконная рекомбинация — термин, используемый для описания негомологичных, аберрантных события рекомбинации, такие как те, которые участвуют в формировании специализированных трансдуцирующие фаги. Механизмы нелегитимной рекомбинации неизвестный.
Транспозоны
Транспозоны представляют собой сегменты ДНК, которые могут перемещаться из одного участка молекулы ДНК в другой.
другие сайты-мишени в той же или другой молекуле ДНК. Процесс
называется транспозицией и происходит по механизму, который не зависит от
генерализованная рекомбинация. Транспозоны являются важными генетическими элементами, потому что
они вызывают мутации, опосредуют геномные перестройки, функционируют как портативные
участки генетической гомологии, приобретают новые гены и вносят свой вклад в их
диссеминация в бактериальных популяциях. Вставка транспозона часто
прерывает линейную последовательность гена и инактивирует его. Транспозоны имеют
главную роль в возникновении делеций, дупликаций и инверсий сегментов ДНК.
а также слияния между репликонами. Транспозоны не являются самовоспроизводящимися генетическими
элементов, однако они должны интегрироваться в другие репликоны, чтобы поддерживать
стабильно в бактериальных геномах.
Вставка транспозона часто
прерывает линейную последовательность гена и инактивирует его. Транспозоны имеют
главную роль в возникновении делеций, дупликаций и инверсий сегментов ДНК.
а также слияния между репликонами. Транспозоны не являются самовоспроизводящимися генетическими
элементов, однако они должны интегрироваться в другие репликоны, чтобы поддерживать
стабильно в бактериальных геномах.
Большинство транспозонов имеют ряд общих черт. Каждый транспозон кодирует
функции, необходимые для его транспозиции, включая фермент транспозазу, который
взаимодействует со специфическими последовательностями на концах транспозона.В течение
при транспозиции короткая последовательность ДНК-мишени удваивается, и транспозон
вставляется между непосредственно повторяющимися целевыми последовательностями. Длина этого
короткая дупликация варьирует, но характерна для каждого транспозона. То
предполагается, что дупликация включает асимметричное расщепление ДНК в мишени.
сайте с последующим синтезом новых комплементарных цепей, соответствующих
области между участками расщепления. Некоторые транспозоны встраиваются практически в любой
целевая последовательность, в то время как другие имеют относительно строгую целевую специфичность.Различают два типа транспозиции. Иссечение транспозона из
донорский сайт с последующей его вставкой в целевой сайт называется нерепликативным
транспозиция. Если транспозон на донорском участке реплицируется и копия
вставляется в целевой сайт, однако этот процесс называется репликативным
транспозиция. Процесс репликативной транспозиции может включать образование
коинтеграт, единая кольцевая молекула ДНК, состоящая из двух соединенных репликонов
с копиями транспозона в чередующейся последовательности.Резолюция
коинтеграция в составные репликоны часто осуществляется с помощью
транспозон-кодируемая резольваза, которая катализирует сайт-специфическую рекомбинацию между
транспозоны. Генерализованная рекомбинация между гомологичными транспозонами может
также приводят к образованию или разрешению коинтегратов. Транспозиция отличается
от сайт-специфической рекомбинации путем дублирования сегмента последовательности-мишени
и с использованием множества различных последовательностей-мишеней для одного донора
последовательность.
Некоторые транспозоны встраиваются практически в любой
целевая последовательность, в то время как другие имеют относительно строгую целевую специфичность.Различают два типа транспозиции. Иссечение транспозона из
донорский сайт с последующей его вставкой в целевой сайт называется нерепликативным
транспозиция. Если транспозон на донорском участке реплицируется и копия
вставляется в целевой сайт, однако этот процесс называется репликативным
транспозиция. Процесс репликативной транспозиции может включать образование
коинтеграт, единая кольцевая молекула ДНК, состоящая из двух соединенных репликонов
с копиями транспозона в чередующейся последовательности.Резолюция
коинтеграция в составные репликоны часто осуществляется с помощью
транспозон-кодируемая резольваза, которая катализирует сайт-специфическую рекомбинацию между
транспозоны. Генерализованная рекомбинация между гомологичными транспозонами может
также приводят к образованию или разрешению коинтегратов. Транспозиция отличается
от сайт-специфической рекомбинации путем дублирования сегмента последовательности-мишени
и с использованием множества различных последовательностей-мишеней для одного донора
последовательность.
Большинство транспозонов у бактерий можно разделить на три основных класса ().Последовательности вставки и связанные с ними
составные транспозоны составляют первый класс. Последовательности вставки самые простые
в структуре и кодируют только функции, необходимые для транспонирования. Известный
последовательности вставки различаются по длине примерно от 780 до 1500 нуклеотидов.
пары, имеют короткие (15-25 пар оснований) инвертированные повторы на концах и не
тесно связаны друг с другом. ДНК между инвертированными концевыми повторами
содержит один (реже два) гена транспозазы и не кодирует резольвазу.Сложные транспозоны имеют длину от 2000 до более 40000
пары нуклеотидов и содержат вставочные последовательности (или близкородственные последовательности)
на каждом конце, обычно в виде перевернутых повторов. Весь сложный элемент может
транспонировать как единое целое. ДНК между концевыми вставками комплекса
транспозоны кодируют несколько функций, которые не являются существенными для транспозиции. У важных с медицинской точки зрения бактерий гены, определяющие прилипание
антигены, токсины или другие факторы вирулентности, или определить устойчивость к одному или
больше антибиотиков, часто располагаются в сложных транспозонах.Известные примеры
сложных транспозонов: Tn 5 и
Tn 10 , определяющие устойчивость к канамицину и
тетрациклин соответственно. Сложные транспозоны, вероятно, эволюционируют
транспозиция гомологичных последовательностей вставок в близлежащие участки ДНК
молекула.
У важных с медицинской точки зрения бактерий гены, определяющие прилипание
антигены, токсины или другие факторы вирулентности, или определить устойчивость к одному или
больше антибиотиков, часто располагаются в сложных транспозонах.Известные примеры
сложных транспозонов: Tn 5 и
Tn 10 , определяющие устойчивость к канамицину и
тетрациклин соответственно. Сложные транспозоны, вероятно, эволюционируют
транспозиция гомологичных последовательностей вставок в близлежащие участки ДНК
молекула.
Рисунок 5-11
Интегрированные репрезентативные транспозоны (жирные линии) в бактериальную хромосому (тонкие линии). Транспозоны являются важными генетическими элементами, поскольку они вызывают мутации, опосредованные геномные перестройки, функционируют как портативные области генетических (более…)
Второй класс транспозонов состоит из высоко гомологичных
Тн Семейство . Эти транспозоны имеют более длинные (от 35 до 40
пара оснований) концевые инвертированные повторы, чем описанные комплексные транспозоны
выше, но в них отсутствуют терминальные последовательности вставки. Все члены семьи
кодируют как функции транспозазы, так и функции резольвазы. Известные примеры из
Tn Семейство транспозонов включает ампициллин
резистентность транспозона Tn 3 и
Tn 1000 (гамма-дельта-транспозон) найден в F
плазмида.Семья TnA занимает важное место в истории медицины.
микробиология. Развитие высокой устойчивости к ампициллину у Haemophilus influenzae и Neisseria
gonorrhoeae в 1970-х годах, что сильно ограничило полезность
ампициллина для лечения гонореи и гемофильной инфекции в районах
где такие штаммы стали преобладать, было вызвано распространением ампициллина
детерминанты устойчивости из транспозонов Tn A в плазмидах
Enterobacteriaceae к плазмидам Haemophilus и Нейссерия .
Все члены семьи
кодируют как функции транспозазы, так и функции резольвазы. Известные примеры из
Tn Семейство транспозонов включает ампициллин
резистентность транспозона Tn 3 и
Tn 1000 (гамма-дельта-транспозон) найден в F
плазмида.Семья TnA занимает важное место в истории медицины.
микробиология. Развитие высокой устойчивости к ампициллину у Haemophilus influenzae и Neisseria
gonorrhoeae в 1970-х годах, что сильно ограничило полезность
ампициллина для лечения гонореи и гемофильной инфекции в районах
где такие штаммы стали преобладать, было вызвано распространением ампициллина
детерминанты устойчивости из транспозонов Tn A в плазмидах
Enterobacteriaceae к плазмидам Haemophilus и Нейссерия .
Третий класс транспозонов состоит из бактериофага Mu и родственных умеренных
фаги. Весь геном фага функционирует как транспозон, и репликация
ДНК фага во время вегетативного роста происходит путем репликативной транспозиции. Интеграция профага может происходить во многих различных местах бактериального
хромосомы и часто вызывает мутации. По этой причине Mu и родственные фаги
иногда называемые фагами-мутаторами.
Интеграция профага может происходить во многих различных местах бактериального
хромосомы и часто вызывает мутации. По этой причине Mu и родственные фаги
иногда называемые фагами-мутаторами.
Четвертый класс транспозонов, обнаруженный у грамположительных бактерий и
представлен Tn 917 , состоит из конъюгативных транспозонов
которые полностью отличаются от транспозонов, описанных выше.То
конъюгативный транспозон не продуцирует дупликацию целевой последовательности
в которую он встраивается, а у грамположительных бактерий штамм-хозяин, несущий
транспозон может действовать как супружеский донор. Бактерии-реципиенты не обязательно
близкие к бактериям-донорам. Транспозон вырезается из
хромосомы донора и передается путем конъюгации реципиенту, где
он случайным образом интегрируется в хромосому. Tn 917 кодирует
резистентность к тетрациклину, но другие более крупные конъюгативные транспозоны могут кодировать
дополнительная устойчивость к антибиотикам.Конъюгативные транспозоны, по-видимому, являются основными
причиной распространения антибиотикорезистентности у грамположительных бактерий.
Некоторые роли транспозонов в эволюции бактерий иллюстрируются рассмотрением
кишечные грамотрицательные бактерии и строение их плазмид. Бактерии
собранные в доантибиотиковую эпоху, содержали много плазмид, но они
обычно отсутствуют детерминанты резистентности. Многие R-плазмиды из современных
клинические изоляты принадлежат к тем же группам несовместимости, что и обнаруженные плазмиды
ранее, но они также определяют устойчивость ко многим антибиотикам.То
тесная связь между их репликонами убедительно свидетельствует о том, что многие
современные плазмиды R произошли от более старых плазмид путем приобретения устойчивости
детерминанты. Некоторые из множественных устойчивых к антибиотикам плазмид имеют индивидуальные
транспозоны с несколькими детерминантами резистентности, другие имеют множественные
транспозоны резистентности расположены в отдельных сайтах, а третьи содержат
сложные гибридные транспозоны устойчивости, образованные путем интеграции одного транспозона
в другой. Поэтапное приобретение детерминант резистентности может привести, в
некоторых случаях к образованию составных транспозонов, кодирующих несколько
детерминанты резистентности. Терапевтическое применение антибиотиков и их инкорпорация
в корма для животных обеспечивают селективные преимущества для бактерий с R-плазмидами,
тогда как конъюгация, трансформация и трансфекция обеспечивают средства для
диссеминация R-плазмид внутри и между видами бактерий. После
плазмида, несущая транспозон, вводится в новую бактерию-хозяина.
транспозон и его детерминанты могут прыгать в хромосому или коренные
плазмиды нового хозяина. Таким образом, стабильность мобилизационной плазмиды в
новый бактериальный хозяин не является существенным для персистенции генетических детерминант
расположен на транспозоне.
Терапевтическое применение антибиотиков и их инкорпорация
в корма для животных обеспечивают селективные преимущества для бактерий с R-плазмидами,
тогда как конъюгация, трансформация и трансфекция обеспечивают средства для
диссеминация R-плазмид внутри и между видами бактерий. После
плазмида, несущая транспозон, вводится в новую бактерию-хозяина.
транспозон и его детерминанты могут прыгать в хромосому или коренные
плазмиды нового хозяина. Таким образом, стабильность мобилизационной плазмиды в
новый бактериальный хозяин не является существенным для персистенции генетических детерминант
расположен на транспозоне.
Рекомбинация ДНК и клонирование генов
Существует множество методов получения гибридных молекул ДНК in vitro (рекомбинантная ДНК)
и дать им характеристику. К таким методам относится выделение специфических генов в гибридах.
репликонов, определяя их нуклеотидные последовательности и создавая мутации в
обозначенные места (сайт-направленный мутагенез). Клон – это популяция
организмы или молекулы, полученные путем бесполого размножения от одного предка. Ген
клонирование — это процесс включения чужеродных генов в репликоны гибридной ДНК.Клонированные гены могут быть экспрессированы в соответствующих клетках-хозяевах, а фенотипы,
они определяют, могут быть проанализированы. Некоторые ключевые концепции, лежащие в основе репрезентативных методов
обобщаются здесь.
Ген
клонирование — это процесс включения чужеродных генов в репликоны гибридной ДНК.Клонированные гены могут быть экспрессированы в соответствующих клетках-хозяевах, а фенотипы,
они определяют, могут быть проанализированы. Некоторые ключевые концепции, лежащие в основе репрезентативных методов
обобщаются здесь.
Первым этапом клонирования генов является получение фрагментов донорской ДНК механическим или
ферментативные методы. Определенные эндонуклеазы рестрикции, относящиеся к классу II,
особенно полезен для получения определенных фрагментов молекул ДНК. Они расщепляются
обе цепи двухцепочечных молекул ДНК в определенных палиндромных последовательностях
(сайты рестрикции), которые обычно имеют длину от четырех до восьми нуклеотидов, и
полученные фрагменты ДНК называются фрагментами рестрикции.Некоторые ограничения
эндонуклеазы расщепляются в совпадающих участках с образованием фрагментов ДНК с тупыми концами и
другие разрезают в шахматном порядке, чтобы создать фрагменты ДНК с короткими,
самокомплементарные, одноцепочечные 5′- или 3′-концы (см. ). Случайная вероятность того, что n
соседние нуклеотиды в цепи ДНК будут соответствовать определенному сайту рестрикции
составляет примерно 1/4 n . Сайты для ферментов, которые распознают уникальные 4, 6 или 8
нуклеотидные мишени, вероятно, встречаются примерно один раз на каждые 256, 4096 или 65 536
нуклеотидов соответственно.Выбирая соответствующие ферменты рестрикции, специфическая ДНК
молекулы, включая бактериальные хромосомы, плазмиды и фаговые геномы, могут быть
переварены в наборы рестрикционных фрагментов, которые имеют соответствующие размеры для конкретных
Приложения.
). Случайная вероятность того, что n
соседние нуклеотиды в цепи ДНК будут соответствовать определенному сайту рестрикции
составляет примерно 1/4 n . Сайты для ферментов, которые распознают уникальные 4, 6 или 8
нуклеотидные мишени, вероятно, встречаются примерно один раз на каждые 256, 4096 или 65 536
нуклеотидов соответственно.Выбирая соответствующие ферменты рестрикции, специфическая ДНК
молекулы, включая бактериальные хромосомы, плазмиды и фаговые геномы, могут быть
переварены в наборы рестрикционных фрагментов, которые имеют соответствующие размеры для конкретных
Приложения.
Таблица 5-3
Особенности репрезентативного класса II Ограничение Эндонуклеазы.
Карта ограничений определяет позиции целевых сайтов для конкретных ограничений эндонуклеаз в молекуле ДНК. Карты рестрикции доступны для многих клонированных ДНК. фрагментов, плазмид и фаговых геномов, а также для всей хромосомы Э.coli и несколько других бактерий.
Вторым этапом клонирования генов является создание гибридных репликонов, состоящих из донорской ДНК. фрагменты и клонирующий вектор ().
Клонирующие векторы представляют собой небольшие плазмидные или фаговые репликоны, имеющие один или несколько
сайты рестрикции, в которые может быть вставлена чужеродная ДНК. Гибридные репликоны
производится с использованием ДНК-лигазы для соединения рестриктированной векторной ДНК с ДНК донора
фрагменты, имеющие совместимые концы, или, альтернативно, синтетические олигонуклеотиды
используются в качестве линкеров для создания совместимости между донорными и векторными молекулами ДНК
с разными концами.Лигирование вектора в гетерогенный набор фрагментов ДНК из
геном донора называется клонированием дробовиком, а сбор рекомбинантной ДНК
молекул, содержащих различные фрагменты, называется геномной библиотекой. Если
специфический фрагмент ДНК доступен, он может быть включен в рекомбинантную
репликон путем прямого клонирования в подходящий вектор, выбранный из большого разнообразия
доступных векторов. Плазмидные и фаговые векторы используются в основном для клонирования небольших
вставки обычно менее 10 kbp.
фрагменты и клонирующий вектор ().
Клонирующие векторы представляют собой небольшие плазмидные или фаговые репликоны, имеющие один или несколько
сайты рестрикции, в которые может быть вставлена чужеродная ДНК. Гибридные репликоны
производится с использованием ДНК-лигазы для соединения рестриктированной векторной ДНК с ДНК донора
фрагменты, имеющие совместимые концы, или, альтернативно, синтетические олигонуклеотиды
используются в качестве линкеров для создания совместимости между донорными и векторными молекулами ДНК
с разными концами.Лигирование вектора в гетерогенный набор фрагментов ДНК из
геном донора называется клонированием дробовиком, а сбор рекомбинантной ДНК
молекул, содержащих различные фрагменты, называется геномной библиотекой. Если
специфический фрагмент ДНК доступен, он может быть включен в рекомбинантную
репликон путем прямого клонирования в подходящий вектор, выбранный из большого разнообразия
доступных векторов. Плазмидные и фаговые векторы используются в основном для клонирования небольших
вставки обычно менее 10 kbp. Примеры векторов более специального назначения включают
космиды, которые представляют собой плазмидные векторы, которые могут быть упакованы в фаговые капсиды (лямбда
космиды принимают вставки размером до 30-40 т.п.н.), а фагемиды, представляющие собой плазмидно-фаговые
гибридные репликоны, которые могут существовать либо в виде плазмид, либо в виде одноцепочечных ДНК-фагов
в разных экспериментальных условиях.Космиды фага P1 могут принимать вставки до
100 т.п.н. и даже более крупные молекулы ДНК могут быть клонированы в искусственных дрожжах.
хромосом (YAC), которые могут стабильно поддерживать вставки размером до 1 Мбп и выше в
размер. Другие специализированные векторы обнаруживают промоторы, сигналы терминации транскрипции,
или другие регуляторные элементы в составе чужеродных ДНК-вставок или, наоборот, обеспечивают
промоторы, с которых может быть инициирована транскрипция клонированных генов.
Примеры векторов более специального назначения включают
космиды, которые представляют собой плазмидные векторы, которые могут быть упакованы в фаговые капсиды (лямбда
космиды принимают вставки размером до 30-40 т.п.н.), а фагемиды, представляющие собой плазмидно-фаговые
гибридные репликоны, которые могут существовать либо в виде плазмид, либо в виде одноцепочечных ДНК-фагов
в разных экспериментальных условиях.Космиды фага P1 могут принимать вставки до
100 т.п.н. и даже более крупные молекулы ДНК могут быть клонированы в искусственных дрожжах.
хромосом (YAC), которые могут стабильно поддерживать вставки размером до 1 Мбп и выше в
размер. Другие специализированные векторы обнаруживают промоторы, сигналы терминации транскрипции,
или другие регуляторные элементы в составе чужеродных ДНК-вставок или, наоборот, обеспечивают
промоторы, с которых может быть инициирована транскрипция клонированных генов.
Рисунок 5-12
Схематическое изображение эксперимента по клонированию генов. Плазмидный вектор клонирования pBR322 имеет размер 4,36 тыс. пар оснований, содержит гены
на устойчивость к ампициллину (amp r ) и к тетрациклину
( tet r ) и имеет только один HindIII
сайт рестрикции, расположенный в (подробнее…)
Плазмидный вектор клонирования pBR322 имеет размер 4,36 тыс. пар оснований, содержит гены
на устойчивость к ампициллину (amp r ) и к тетрациклину
( tet r ) и имеет только один HindIII
сайт рестрикции, расположенный в (подробнее…)
Заключительным этапом клонирования генов является введение гибридных репликонов в соответствующие
реципиентные клетки и проверить их на экспрессию интересующих донорских генов. прокариотический
клетки (включая бактерии) или эукариотические клетки (включая дрожжевые, животные или растительные
клетки) могут быть использованы в качестве реципиентов, но они различаются по своим
допустимость для конкретных репликонов, транскрипционные сигналы, которые они
распознавать, и посттрансляционные модификации структуры белка, которые они
может выполнить.Молекулы рекомбинантной ДНК, полученные in vitro, могут быть введены
непосредственно в клетки-реципиенты путем трансформации или трансфекции. Кроме того, клоны
космидные или фаговые векторы могут быть упакованы в фаговые оболочки и введены в
восприимчивые клетки-реципиенты путем трансдукции. Используя специализированные векторы (челночные
векторов), которые могут реплицироваться во многих типах клеток, гены любого организма могут быть
клонировали и манипулировали в удобной бактериальной системе, а затем
повторно вводили в клетки исходного организма для анализа в их естественном
окружающая обстановка.
Используя специализированные векторы (челночные
векторов), которые могут реплицироваться во многих типах клеток, гены любого организма могут быть
клонировали и манипулировали в удобной бактериальной системе, а затем
повторно вводили в клетки исходного организма для анализа в их естественном
окружающая обстановка.
Многие методы доступны для идентификации бактерий, содержащих рекомбинантную ДНК
молекулы. Большинство клонирующих векторов имеют гены признаков, которые могут быть положительно
выбранных, таких как устойчивость к антибиотикам. Кроме того, часто можно
ввести чужеродную ДНК в клонирующий вектор в сайт, который инактивирует
несущественная, но легко узнаваемая векторная функция. Если оба эти условия
выполнены, можно отобрать бактерии, содержащие рекомбинантные молекулы, и
легко отличить от бактерий, содержащих только переносчик.Бактерии в
геномную библиотеку, содержащую конкретный клонированный ген, можно идентифицировать с помощью
биохимические или иммунологические методы для проверки желаемого генного продукта. В качестве альтернативы интересующий клонированный ген можно обнаружить непосредственно с помощью нуклеиновых
методы кислотной гибридизации, при условии, что имеется специфический ДНК- или РНК-зонд.
Поскольку вставка чужеродной ДНК в клонирующий вектор в соответствующий сайт
не инактивировать свою способность к репликации в соответствующих клетках-реципиентах, гибридных
интересующие репликоны могут быть амплифицированы путем репликации, а рекомбинантная ДНК
молекулы или их генные продукты могут быть очищены и изучены.Способность очищать
специфические молекулы ДНК позволили разработать ферментативные и химические методы
для определения их нуклеотидных последовательностей, а также современные методы введения
мутации в определенных сайтах в клонированных генах основаны на знании их ограничения
карты или последовательности нуклеотидов.
В качестве альтернативы интересующий клонированный ген можно обнаружить непосредственно с помощью нуклеиновых
методы кислотной гибридизации, при условии, что имеется специфический ДНК- или РНК-зонд.
Поскольку вставка чужеродной ДНК в клонирующий вектор в соответствующий сайт
не инактивировать свою способность к репликации в соответствующих клетках-реципиентах, гибридных
интересующие репликоны могут быть амплифицированы путем репликации, а рекомбинантная ДНК
молекулы или их генные продукты могут быть очищены и изучены.Способность очищать
специфические молекулы ДНК позволили разработать ферментативные и химические методы
для определения их нуклеотидных последовательностей, а также современные методы введения
мутации в определенных сайтах в клонированных генах основаны на знании их ограничения
карты или последовательности нуклеотидов.
Методы рекомбинантной ДНК позволяют клонировать специфические фрагменты ДНК из любого
источник в векторы, которые можно изучать на хорошо охарактеризованных бактериях, в
эукариотических клеток или in vitro . Применение клонирования ДНК
быстро расширяется во всех областях биологии и медицины. В медицинской генетике такие
Область применения варьируется от пренатальной диагностики наследственных заболеваний человека до
характеристика онкогенов и их роли в канцерогенезе. Фармацевтическая
приложения включают крупномасштабное производство из клонированных человеческих генов биологических
продукты с терапевтической ценностью, такие как полипептидные гормоны, интерлейкины и
ферменты. Приложения в общественном здравоохранении и лабораторной медицине включают разработку
вакцин для предотвращения специфических инфекций и зондов для диагностики специфических
инфекций путем гибридизации нуклеиновых кислот или полимеразной цепной реакции (ПЦР).То
последний процесс использует олигонуклеотидные праймеры и ДНК-полимеразу для амплификации специфических
целевые последовательности ДНК во время нескольких циклов синтеза in vitro, что делает его
можно обнаружить редкие последовательности ДНК-мишеней в клинических образцах с большим
чувствительность.
Применение клонирования ДНК
быстро расширяется во всех областях биологии и медицины. В медицинской генетике такие
Область применения варьируется от пренатальной диагностики наследственных заболеваний человека до
характеристика онкогенов и их роли в канцерогенезе. Фармацевтическая
приложения включают крупномасштабное производство из клонированных человеческих генов биологических
продукты с терапевтической ценностью, такие как полипептидные гормоны, интерлейкины и
ферменты. Приложения в общественном здравоохранении и лабораторной медицине включают разработку
вакцин для предотвращения специфических инфекций и зондов для диагностики специфических
инфекций путем гибридизации нуклеиновых кислот или полимеразной цепной реакции (ПЦР).То
последний процесс использует олигонуклеотидные праймеры и ДНК-полимеразу для амплификации специфических
целевые последовательности ДНК во время нескольких циклов синтеза in vitro, что делает его
можно обнаружить редкие последовательности ДНК-мишеней в клинических образцах с большим
чувствительность.
Регуляция экспрессии генов
Фенотипические свойства бактерий определяются их генотипами и ростом условия. Для бактерий в чистой культуре изменение условий роста часто приводит к в предсказуемой физиологической адаптации у всех представителей популяции.Как правило, основные генные продукты производятся в количествах, обеспечивающих быстрый рост данная среда, и продукты, требуемые при особых обстоятельствах, производятся только когда они нужны.
Физиологические адаптации часто связаны с изменениями метаболической активности. Поток метаболитов по определенным биохимическим путям можно контролировать. как за счет регуляции синтеза специфических ферментов, так и за счет изменения активности существующих ферментов. Механизмы, которые регулируют экспрессию генов, влияя на здесь обсуждаются синтезы специфических генных продуктов.
Специфическая регуляция включает ген или группу генов, участвующих в определенном
метаболический процесс. Индукция и репрессия позволяют бактериям регулировать производство
специфических генных продуктов в ответ на соответствующие сигналы. Обычно катаболический
ферменты индуцируются, когда субстрат для пути присутствует в ростовой
среде, а ферменты биосинтеза репрессируются продуктом пути.
Ферменты, участвующие в одном биохимическом пути, часто занимают соседние
позиции на бактериальной хромосоме и координированно индуцируются или репрессируются.Они образуют оперон, группу смежных генов, которая транскрибируется как единый
единица и транслируется для производства соответствующих генных продуктов. Организация в
оперон является важной стратегией координированной регуляции экспрессии генов.
в бактериях. Опероны, которые могут быть индуцированы или репрессированы, контролируются связыванием
специфические регуляторные белки к определенным последовательностям нуклеотидов, которые функционируют как
регуляторные участки оперона. Сравнение аминокислотных последовательностей многих
этих различных регуляторных белков показали, что они могут быть сгруппированы вместе.
на семейства регуляторов (т.г. семейство белков lysR), которые, возможно, эволюционировали
от общих предковых генов.
Обычно катаболический
ферменты индуцируются, когда субстрат для пути присутствует в ростовой
среде, а ферменты биосинтеза репрессируются продуктом пути.
Ферменты, участвующие в одном биохимическом пути, часто занимают соседние
позиции на бактериальной хромосоме и координированно индуцируются или репрессируются.Они образуют оперон, группу смежных генов, которая транскрибируется как единый
единица и транслируется для производства соответствующих генных продуктов. Организация в
оперон является важной стратегией координированной регуляции экспрессии генов.
в бактериях. Опероны, которые могут быть индуцированы или репрессированы, контролируются связыванием
специфические регуляторные белки к определенным последовательностям нуклеотидов, которые функционируют как
регуляторные участки оперона. Сравнение аминокислотных последовательностей многих
этих различных регуляторных белков показали, что они могут быть сгруппированы вместе.
на семейства регуляторов (т.г. семейство белков lysR), которые, возможно, эволюционировали
от общих предковых генов. Члены семейства lysR включают регуляторы таких
разнообразные явления, такие как метаболизм лизина, цистеина и метионина в E.
coli и подавление железа у V. cholerae .
Члены семейства lysR включают регуляторы таких
разнообразные явления, такие как метаболизм лизина, цистеина и метионина в E.
coli и подавление железа у V. cholerae .
Глобальная регуляция одновременно изменяет экспрессию группы генов и оперонов,
вместе называемые регулоном, которые контролируются одним и тем же регуляторным сигналом.
Глобальное регулирование определяет реакцию бактерий на основные питательные вещества, такие как
углерод, азот или фосфат, реакции на стрессы, такие как повреждение ДНК или тепло
шок, синтез возбудителями специфических факторов вирулентности при росте в
животных-хозяев.Количество определенного белка в бактериальной клетке может варьироваться.
от нуля до многих тысяч молекул. Этот широкий диапазон часто определяется
комбинированное действие нескольких регуляторных механизмов, влияющих на экспрессию
соответствующий структурный ген. Регулирование достигается путем определения того, как часто
ген транскрибируется в функциональную мРНК, насколько эффективно транслируется мРНК
в белок, как быстро расщепляется мРНК, как быстро белковый продукт
переворачивается, и может ли активность белкового продукта быть изменена
аллостерические эффекты или ковалентные модификации.
мРНК как единицы транскрипции
Экспрессия генов начинается с ДНК-зависимой РНК-полимеразы (РНК-полимеразы) катализирует транскрипцию специфической мРНК с одной цепи ДНК-матрицы. Связывание РНК-полимеразы с ДНК происходит в определенных местах, называемых промоторами. транскрипция начинается рядом с промотором. Сильные промоутеры могут взаимодействовать эффективно с РНК-полимеразой и инициировать транскрипцию с высокой скоростью; слабый промоторы инициируют транскрипцию с низкой скоростью. В любом случае мРНК синтезируется от 5′-конца к 3′-концу на примерно с постоянной скоростью до тех пор, пока РНК-полимераза не распознает другой специфический сайт назвал терминатором.Затем РНК-полимераза диссоциирует от матрицы и транскрипция мРНК завершается.
Отдельные молекулы мРНК могут кодировать один или несколько полипептидов. Транскрипция оперон производит полицистронную мРНК, которая кодирует несколько полипептидов. Трансляция полицистронных мРНК приводит к координированному синтезу кодируемых полипептиды, но каждый полипептид синтезируется как отдельная молекула. А специфический сайт связывания рибосомы расположен непосредственно перед началом каждой кодирующая последовательность молекулы мРНК.
Информационная РНК в бактериях быстро разрушается со средним периодом полураспада несколько минут, в отличие от тРНК и рРНК, которые гораздо более стабильны. Хотя мРНК составляют примерно половину вновь синтезированной РНК, они представляют лишь небольшая часть всей РНК. Короткий период полураспада мРНК важные последствия для экспрессии генов. Если синтез конкретной мРНК предотвращается, продукция соответствующих полипептидов быстро снижается.
Контроль экспрессии генов осуществляется путем регуляции одного или нескольких этапов в путь от матрицы ДНК к продукту активного гена.Одновременный регуляция на нескольких уровнях позволяет лучше контролировать экспрессию генов, чем было бы возможно при наличии единого регулирующего механизма. Самый распространенный способ Регуляция экспрессии генов у бактерий заключается в контроле производства специфических мРНК. Поскольку скорость удлинения молекулы РНК примерно равна постоянными, основными факторами, контролирующими синтез мРНК, являются скорость инициирование и вероятность того, что полная стенограмма будет произведено.
Регуляция инициации транскрипции
Некоторые мРНК у бактерий синтезируются с постоянной скоростью, что приводит к конститутивная продукция кодируемых полипептидов.Количество конкретных мРНК и полипептиды, полученные из разных конститутивных генов, сильно различаются. тем не менее, и часто отражают различия в силе промоторов для тех, гены.
Транскрипция многих оперонов регулируется в ответ на изменение окружающей среды условия. Промоторы определяют максимальную скорость инициации транскрипции. для таких оперонов, но регуляторные белки участвуют в контроле транскрипция. Нуклеотидные последовательности в оперонах, к которым относятся специфические регуляторные белки связывания называются регуляторными сайтами или операторами.Операторы и промоутеры расположены близко друг к другу внутри оперонов и могут иметь перекрывающуюся ДНК последовательности. Связывание регуляторных белков с операторами может (положительная регуляция) или уменьшение (отрицательная регуляция) частоты инициация транскрипции. Белки, выполняющие функцию негативных регуляторов, обычно называют репрессорами. Поскольку регуляторные белки могут диффундировать через цитоплазме, структурные гены регуляторных белков не обязательно должны быть связаны к целевым оперонам.
Способность ощущать присутствие или отсутствие определенных соединений и изменять скорость синтеза соответствующих генных продуктов играет центральную роль в контроле экспрессия генов. Регуляторные белки предлагают одно из решений этой проблемы связь стимул-реакция. Многие регуляторные белки бифункциональны и связывают не только к соответствующим операторам, но и к специфическим эффекторам, которые небольшие молекулы, такие как определенные сахара, аминокислоты и другие метаболиты. Кроме того, регуляторные белки являются аллостерическими, что означает, что они могут существовать в различные конформации, которые проявляют различную аффинность связывания для своих родственные операторы и эффекторы.Достаточная концентрация эффектора способствует образование регуляторного белково-эффекторного комплекса, обладающего либо высоким, либо низкая аффинность к оператору в каждом конкретном случае. В отрицательно регулируемых систем эффектор функционирует как корепрессор, если регуляторная белок-эффекторный комплекс является активным репрессором, а эффектор действует как индуктор и вызывает дерепрессию, если свободный регуляторный белок является активным репрессор. И наоборот, в положительно регулируемых системах эффектор стимулирует экспрессия оперона, если регуляторный белок-эффекторный комплекс является положительный регулятор, а эффектор ингибирует экспрессию оперона, если свободный регуляторный белок является положительным регулятором.
Лактозный (lac) оперон E. coli является примером индуцируемый, отрицательно регулируемый оперон (). Ген lacI кодирует репрессор, который связывается с оператором lac и предотвращает транскрипцию с lac промоутер. Структурный ген этого репрессора отделены от lac оперона, а репрессор конститутивно синтезируется в низкая ставка. Когда индуктор связывается с lac-репрессором, комплекс не может связываться с оператора и не может предотвратить связывание РНК-полимеразы с промотором.При благоприятных других условиях экспрессируется оперон lac , что приводит к синтезу β-галактозидазы, β-галактозида пермеазы и β-галактозидтрансацетилазы. лак оперон может индуцироваться лактозой или структурно родственными соединениями, такими как изопропил-b-D-тиогалактозид (IPTG). ИПТГ называют безвозмездным индуктором потому что он индуцирует lac оперон, но не является субстратом для β-галактозидаза. Негативная регуляция также имеет место во многих биосинтетических опероны в E.палочка . В таких оперонах продукт биосинтетический путь функционирует как эффектор негативной регуляторной система.
Рисунок 5-13
Регуляция оперона lac в E. палочка . Структурные гены lacZ , lacY и lacA код для β-галактозидазы, β-галактозидпермеаза и β-галактозидтрансацетилаза, соответственно. Физиологическая роль lacA заключается в следующем. неизвестный. Репрессор lac (подробнее…)
Оперон арабинозы ( ara ) в E. coli является одновременно позитивно и негативно регулируется. В присутствии арабинозы регуляторный белок стимулирует транскрипцию оперона ara. В отсутствие арабинозы, однако регуляторный белок репрессирует оперон ara .
Опероны часто контролируются более чем одним механизмом. Когда Э. coli выращивают в среде, содержащей глюкозу и альтернативный источник углерода, такой как лактоза или арабиноза, индукция lac или ara оперон и использование лактоза или арабиноза задерживаются до тех пор, пока не будет израсходована глюкоза.Этот явление называется диауксическим ростом. Неспособность индуцировать lac или ara оперон в присутствии глюкозы является примером катаболитной репрессии. лак и ara опероны положительно регулируются цикло-3′,5′-аденозинмонофосфат (цАМФ) и белок-активатор гена катаболита (CAP) (продукт гена crp). То Комплекс cAMP-CAP взаимодействует с сайтами связывания CAP в регуляторных областях некоторые опероны, включая опероны lac и ara, и стимулирует транскрипцию от соответствующих промоутеров.Уровень внутриклеточного цАМФ у E. coli высокая в период роста в отсутствие глюкозы и низкая при росте в присутствии глюкозы. Катаболитная репрессия обусловлена, следовательно, к отсутствию активации цАМФ-зависимых оперонов, когда бактерии выращивают в присутствии глюкозы или некоторых других быстро метаболизируемых углеродных источники.
Регуляция терминации транскрипции
Аттенуация – это механизм регуляции оперонов путем терминации транскрипции мРНК преждевременно.Аттенуация характерна для биосинтетических оперонов, включая trp , гистидин ( его ), треонин ( через ), изолейцин-валин ( ilv ) и фенилаланиновые (phe) опероны. Оперон trp в E. coli контролируется как репрессией, так и аттенуацией. в наличие избытка триптофана, инициация транскрипции с Промотор trp репрессирован. Кроме того, однако, те транскрипты, инициированные промотором trp , обычно терминируется перед любым из структурных генов trp оперона транскрибируются.Концентрация внутриклеточный триптофан, необходимый для поддержания репрессии, превышает необходимый для затухание. Такой двойной контроль позволяет клетке точно регулировать экспрессию оперона trp в ответ на снижение концентрации триптофан.
Вторичная структура мРНК играет важную роль в механизме затухание. Все мРНК имеют лидерную последовательность между началом транскрипции сайт и начало кодирующей последовательности для первого структурного гена.За опероны биосинтеза аминокислот, подверженные аттенуации, лидер мРНК последовательность имеет две отличительные особенности. Он кодирует короткий пептид, содержащий аминокислота, вырабатываемая регулируемым путем, и она может образовывать альтернативные, взаимно несовместимые двухцепочечные структуры РНК, участвующие в регламентные мероприятия. Например, пептид, кодируемый trp Лидерная последовательность мРНК содержит два соседних остатка триптофана, а пептид кодируется , его лидерная последовательность мРНК имеет серию из семи последовательные остатки гистидина.показывает оперон trp и иллюстрирует альтернативный вторичный структуры в лидерной последовательности мРНК trp . Есть три возможные вторичные структуры для этой области, называемые местом паузы (сегменты 1+2), антитерминатор (сегменты 2+3) и аттенюатор (сегменты 3+4). Сегмент 1 участка паузы перекрывается с кодировкой участок для пептида trpL . Какие вторичные структуры формируется зависит от эффективности трансляции trpL пептид.Когда сегменты 1 и 2 транскрибируются, они немедленно отжигаются и вызвать временную остановку РНК-полимеразы. Последующее инициирование трансляция пептида trpL нарушает сайт паузы и позволяет РНК-полимеразе продолжать транскрипцию. Если присутствует триптофан, транскрипция сегментов 3 и 4 и формирование аттенюаторной структуры происходит, когда рибосома блокирует сегмент 2, вызывая РНК-полимеразу прекратить транскрипцию. Однако при дефиците триптофана триптофанил-тРНК также недостаточен, и рибосома останавливается на триптофановых кодонах в сегменте 1.Это позволяет сегменту 2 отжигаться с вновь синтезированным сегментом 3, чтобы сформировать антитерминатора, тем самым делая сегмент 3 недоступным для отжига с сегментом 4. Таким образом предотвращается образование аттенюатора, и РНК-полимераза транскрибирует весь оперон trp . Таким образом, истощение триптофан (фактически источник триптофанил-тРНК) связан с регуляцией транскрипции биосинтетического оперона триптофана.
Рисунок 5-14
Регуляция оперона trp в E.палочка . Организация оперона trp показана на рис. вершина фигуры. Пять структурных генов trpE , трпД , трпК , trpB и trpA кодируют ферменты которые катализируют терминальную последовательность реакций в триптофане формирование. (подробнее…)
В E. coli транскрипция и трансляция функционально в паре. Бессмысленные мутации, вызывающие преждевременное прекращение трансляции часто вызывают снижение транскрипции более дистальных генов в том же опероне.Это явление называется полярностью. Рибосомы обычно инициируют трансляцию растущая молекула мРНК до завершения транскрипции, и такая трансляция маскирует сайты, которые в противном случае вызвали бы терминацию РНК-полимеразы транскрипция. Преждевременная терминация трансляции бессмысленным кодоном отделяет рибосомы от мРНК и позволяет РНК-полимеразе взаимодействовать с немаскированными сайтами терминации транскрипции.
В некоторых биологических системах, включая фаг лямбда, антитерминация используется как положительный регуляторный механизм для контроля экспрессии генов.Незамедлительно после инфекции E. coli лямбда, РНК-полимераза связывается с двумя промоторы в ДНК лямбда и инициирует расходящиеся первичные транскрипты, которые заканчиваются на определенных участках генома лямбда. Белок, кодируемый одним из первичные транскрипты взаимодействуют с РНК-полимеразой и позволяют ей продолжать транскрипция через первичные сайты терминации, тем самым экспрессируя второй набор лямбда-генов. Один из продуктов, кодируемых вторичным транскриптом блокирует терминацию другой мРНК и активирует экспрессию третьего набора гены.Таким образом, антитерминация играет ключевую роль в управлении каскадом экспрессия генов при литическом росте фага лямбда. Антитерминация также участвует в регуляции оперонов рРНК E. coli .
Регуляция трансляции
Сайт связывания рибосомы на мРНК комплементарен последовательности в конце 3′-конец 16S рРНК. Взаимодействие между этими последовательностями облегчает образование комплекса инициации синтеза белка. И степень гомология с 16S рРНК и расстояние между сайтом связывания рибосомы и инициирующий кодон влияет на эффективность инициации трансляции.Использование кодонов в мРНК также влияет на эффективность трансляции. РНК-мессенджеры для белков, которые требуются в больших количествах, как правило, используют кодоны, которые транслируются наиболее многочисленные виды тРНК, верно и обратное.
Контроль трансляции важен для регуляции синтеза рибосомных белки. Производство рибосом сопряжено с высокими метаболическими затратами для бактерий. а при высоких скоростях роста рибосомы могут составлять почти половину клетки масса. Большинство рибосомных белков и рРНК собраны в рибосомы. и пул свободных рибосомных субъединиц очень мал.Гены для рибосом белки организованы в несколько оперонов. Некоторые из свободных рибосом Белки непосредственно ингибируют трансляцию полицистронных мРНК, кодирующих их, тем самым обеспечивая баланс синтеза рибосомных белков с требования к их использованию.
Регулоны и белки, передающие сигналы
Регулон представляет собой группу генов или оперонов, контролируемых общим регулятором. Там Есть несколько преимуществ в том, чтобы поставить разные опероны под контроль одного и того же регулятор.Это позволяет сочетать восприятие одного стимула с экспрессия большого количества генов, которые могут быть необходимы для соответствующего ответ, и это устраняет потребность в координированно регулируемых генах быть сцепленным с бактериальной хромосомой. Раздражитель, на который действует регулон отвечает может быть внутриклеточным компонентом или сигналом окружающей среды. Отдельные опероны также могут регулироваться несколькими различными механизмы и выражены в условиях, отличных от тех, которые влияют на целый регулон.
Более 40 различных регулонов были идентифицированы в E. coli . Конкретные примеры регулонов, которые реагируют на внутриклеточные компоненты, включают регулон cAMP-CAP, описанный ранее, и регулоны, контролируемые строгий ответ и ответ SOS. Когда рибосомы сталкиваются с незаряженной тРНК молекул во время синтеза белка активируется строгая реакция и приводит к быстрому прекращению синтеза рРНК. Новый нуклеотид под названием гуанозин-3′-дифосфат-5′-дифосфат (ppGpp) накапливается при аминокислотном голодании.По-видимому, ppGpp, продуцируемый бездействующими рибосомами, быть посредником жесткой реакции, но точный механизм, вызывающий ингибирование синтеза рРНК неизвестно. Ответ SOS связан с повреждение ДНК и включает индукцию более 20 генов, участвующих в нескольких Пути репарации ДНК. Продукт гена recA обнаруживает ингибирование ДНК синтез и инициирует события, ведущие к протеолитическому расщеплению и инактивации репрессора пути SOS, кодируемого lexA ген.
Некоторые регулоны индуцируются специфическими стимулами окружающей среды, такими как питательные вещества ограничения или осмотического стресса. Часто опероны из более чем одного регулона могут быть индуцируется, и термин «стимулон» используется для описания набора генов, индуцированный. Как правило, бактерии ощущают такие условия окружающей среды двумя компонентные системы. Первый компонент представляет собой трансмембранный белок с внеклеточный и внутриклеточный домены. Его внеклеточный домен обнаруживает стимул окружающей среды, и его цитоплазматический домен передает сигнал.То второй компонент представляет собой бифункциональный цитоплазматический белок. У него есть домен получателя взаимодействующий с передающим модулем первого компонента, а также эффекторный домен, контролирующий экспрессию соответствующего регулона. То модули передатчика и приемника двухкомпонентных систем регулирования от широкий спектр регулонов генетически связан и имеет гомологию аминокислот. Сигнально-детектирующие и эффекторные домены белков из разных Однако регулоны различаются и определяют обнаруживаемый сигнал и опероны, которые активируются или репрессируются в ответ на этот сигнал.
Глобальное регулирование играет важную роль в физиологии патогенных бактерий. Например, Vibrio cholerae и Bordetella pertussis экспрессируют многие детерминанты вирулентности под управление системами преобразования сигналов, относящимися к двухкомпонентным системы, описанные выше. Экспрессия белков, необходимых для инвазивного фенотип контролируется температурой у шигелл . Yersinia enterocolitica чувствует как окружающую среду, температуры и концентрации ионов кальция и соединяет эти сигналы с экспрессия генов и клеточная локализация генных продуктов, которые подходит для внутриклеточной или внеклеточной среды.В тканях хозяина концентрация свободного железа чрезвычайно низка, и большинство патогенных бактерий имеют системы транспорта железа с высоким сродством, которые индуцируются при низком уровне железа. условия. Синтез дифтерийного токсина С. diphtheriae , шига-токсин от Shigella dysenteriae , экзотоксин А Pseudomonas aeruginosa и другие специфические белков у многих патогенных бактерий индуцируется в условиях дефицита железа. рост. Эти примеры иллюстрируют, как факторы окружающей среды могут регулировать экспрессия генов вирулентности у патогенных бактерий.
Завещания, трасты и поместья Блог профессора
Мировые соглашения могут быть выгодны всем сторонам. Соглашения об урегулировании могут «помочь снизить судебные издержки, облегчить разрешение споров [и] привести стороны к общему пониманию».
Хотя соглашения об урегулировании могут быть выгодными, они сопряжены с определенным риском. «В деле Austin Trust Co. v. Houren соглашение содержало формулировку, запрещающую сторонам предъявлять претензии в будущем.»
Ниже приведена краткая информация по делу:
После смерти отца владельцы наследства осознали, что их распределение будет отложено до тех пор, пока не будет подана федеральная налоговая декларация. Стремясь ускорить процесс распределения, бенефициары заключили соглашение о семейном урегулировании («FSA») со всеми заинтересованными сторонами. FSA обсуждался сторонами, которые признали, что их либо представлял адвокат, либо они сознательно предпочли не быть представленными адвокатом.
FSA содержал выпуск, в котором, среди прочего, были сняты все требования о нарушении фидуциарных обязанностей. Точная формулировка в выпусках соглашения гласит: «любая и вся ответственность, вытекающая из любых и всех Претензий», включая «претензии в любой форме единоличного соучастия, одновременной, грубой или другой небрежности, неправомерного влияния, принуждения, нарушения фидуциарных обязательств». обязанность или другое неправомерное поведение» и определила «охватываемую деятельность» как включающую «(1) создание, эксплуатацию, управление или администрирование Имущества,… или Трастов, (2) распределение любого имущества или активов… Имущество… или Трасты, (3) любые действия, предпринятые (или не предпринятые) на основании настоящего Соглашения или фактов, перечисленных в статье I», (4) «любые Претензии, связанные с, основанные или явные в Раскрытие информации» и (5) «любые Претензии, связанные с фактами, изложенными в Статье I, или основанные на них, или подтвержденные фактами.
FSA было подписано 10 июня 2015 года. В начале 2016 года исполнитель наследства подал федеральную налоговую декларацию, в которой предполагаемый долг в размере 37 миллионов долларов не был указан ни в качестве актива, ни в качестве обязательства. Austin Trust направил письмо с требованием о погашении предполагаемого долга, которое исполнитель отклонил. Austin Trust заявил о нарушении фидуциарной обязанности, а исполнитель утверждал, что эта обязанность была снята. Суд первой инстанции согласился, и последовала апелляция.
В Остин Ко., суд должен был определить, действительно ли соглашение об освобождении от ответственности, прежде чем рассматривать вопрос о том, нарушил ли исполнитель фидуциарную обязанность. Суд перечислил шесть факторов, которые он учитывал при принятии решения об утверждении мирового соглашения. Суд определил, что освобождение было действительным, и ему не нужно было решать, была ли нарушена фидуциарная обязанность.
Хотя мировые соглашения могут быть выгодными, стороны должны проявлять большую осторожность и осмотрительность перед заключением мирового соглашения.
См. Соглашения об освобождении от ответственности и семейном урегулировании споров о доверительном управлении и имуществе , Закон Фримена, 2021 г.
Особая благодарность Джиму Хиллхаусу (Professional Legal Marketing (PLM, Inc.)) за то, что он обратил мое внимание на эту статью.
15 января 2022 г. в Управление недвижимостью, Планирование недвижимости — В целом, Трасты | Постоянная ссылка | Комментарии (0)
Сельскохозяйственная льгота по налогу на наследство
Сельскохозяйственная недвижимость
Вы можете передать часть сельскохозяйственной собственности без уплаты налога на наследство либо при жизни, либо по своему завещанию.
Сельскохозяйственная собственность, подпадающая под действие сельскохозяйственной помощи, представляет собой землю или пастбище, которые используются для выращивания сельскохозяйственных культур или интенсивного разведения животных. Сюда также входят:
- выращивание сельскохозяйственных культур
- конезаводы для разведения и выращивания лошадей и выпаса скота
- деревьев, которые сажают и собирают не реже одного раза в 10 лет (поросль с коротким оборотом)
- земель, которые в настоящее время не обрабатываются в соответствии со Схемой среды обитания
- земли, которые в настоящее время не обрабатываются по схеме севооборота
- стоимость доли молока, связанная с землей
- некоторые сельскохозяйственные акции и ценные бумаги
- хозяйственные постройки, фермерские коттеджи и фермерские дома
На них не распространяется сельскохозяйственная помощь:
- сельскохозяйственное оборудование и машины
- заброшенных зданий
- собранный урожай
- домашний скот
- недвижимость, подлежащая обязательному договору купли-продажи
Местоположение
Собственность может быть занята владельцем или сдана в аренду, но она должна быть частью действующей фермы в:
- Великобритания
- Нормандские острова
- Остров Мэн
- Европейская экономическая зона
Период владения или занятия
Имущество должно находиться в собственности и использоваться для сельскохозяйственных целей непосредственно перед его передачей для:
- 2 года, если занято владельцем, компанией, контролируемой им, или их супругом или гражданским партнером
- 7 лет, если занято кем-то другим
Имущество, полученное путем передачи
Если лицо, совершающее передачу, первоначально унаследовало имущество от кого-либо, кроме супруга или гражданского партнера, период владения исчисляется с даты этой первой смерти.
Если имущество первоначально было приобретено путем передачи, также должны быть соблюдены следующие условия:
- недвижимость должна иметь право на сельскохозяйственную помощь на дату первой передачи
- имущество должно быть занято в сельскохозяйственных целях лицом, осуществляющим более позднюю передачу, или личными представителями лица, от которого первоначально исходило имущество
- имущество должно претендовать на освобождение от ответственности за исключением тестов на занятие и владение
- один из переносов должен произойти после смерти
Фермерские дома и коттеджи
Здания должны иметь характер и размеры, соответствующие осуществляемой сельскохозяйственной деятельности.Имущество оценивается так, как если бы оно могло использоваться только в сельскохозяйственных целях. Любая стоимость сверх этой «сельскохозяйственной стоимости», например, рыночная цена загородного дома, не соответствует требованиям для сельскохозяйственной помощи.
Коттедж или фермерский дом должен быть занят кем-то, занятым в сельском хозяйстве, или:
- работник фермы на пенсии
- супруга или гражданский партнер умершего сельскохозяйственного работника
Они должны занимать недвижимость как:
- арендатор по договору аренды, предоставленному в рамках их прежнего трудового договора
- защищенный арендатор с законными правами
Ставки помощи сельскому хозяйству
Сельскохозяйственная помощь выплачивается в размере 100%, если:
- лицо, которому принадлежала земля, обрабатывало ее самостоятельно
- земля использовалась кем-то другим на основании краткосрочной лицензии на выпас скота
- он был сдан в аренду, которая началась 1 сентября 1995 года или позже
Имущество, которое принадлежало до 10 марта 1981 г., может претендовать на 100% освобождение, если:
- он подпадал бы под действие Приложения 8 Закона о финансах 1975 года, если бы он был передан до этой даты
- лицо, которому оно принадлежало, не имело права на свободное владение между этой датой и датой текущей передачи
В любом другом случае компенсация выплачивается по более низкой ставке 50%.
Заложенное имущество
Вычтите непогашенные ипотечные кредиты или любые другие обеспеченные обязательства по собственности перед расчетом сельскохозяйственной помощи.
Сельскохозяйственные акции и ценные бумаги
Некоторые акции и ценные бумаги компаний имеют право на сельскохозяйственную помощь, если их стоимость:
- передал умершему контроль над компанией на момент смерти
- происходит от сельскохозяйственной собственности, которая является частью активов компании
Узнайте больше о правилах продажи связанного имущества в течение 3 лет после смерти.
Дарение сельскохозяйственной собственности
Если подарок (или часть подарка) подпадал под действие программы помощи сельскому хозяйству на момент его совершения, он все равно будет соответствовать критериям, если он:
- хранился у лица, получившего его, до своей смерти или смерти лица, сделавшего подарок
- является сельскохозяйственной собственностью, которая использовалась в сельскохозяйственных целях с момента дарения
Если лицо, получившее подарок, умирает раньше лица, сделавшего подарок, условия освобождения должны быть соблюдены как в день вручения подарка, так и в момент его смерти.
Если лицо, получающее подарок, продает его до смерти лица, делающего подарок, компании в обмен на акции этой компании, то эти акции рассматриваются как первоначальное имущество.
Любые дополнительные акции, полученные в результате реорганизации или предложения о поглощении, рассматриваются как часть первоначального подарка.
Замена сельскохозяйственного имущества
Если сельскохозяйственная собственность, подпадающая под действие сельскохозяйственной помощи, заменяется другой сельскохозяйственной собственностью, которая также подпадает под льготу (кроме требования о минимальном сроке владения), то должны быть соблюдены все следующие условия, если исходная собственность была подарена:
- вся выручка от продажи должна быть использована для покупки нового имущества
- купля-продажа должна быть «сделкой на рыночных условиях», в которой покупатели и продавцы действуют независимо и не имеют отношения друг к другу
- продажа и покупка должны состояться в течение 3 лет друг от друга
Первоначальная и замещающая собственность вместе должны удовлетворять условиям, указанным в разделе «Подарки сельскохозяйственной собственности».
Если сельскохозяйственное имущество находилось в собственности человека на момент его смерти и заменило другое сельскохозяйственное имущество, оно все еще может претендовать на сельскохозяйственную помощь, если первоначальное и замененное имущество:
- был занят владельцем в сельскохозяйственных целях в течение не менее 2 лет в течение 5 лет непосредственно перед смертью
- находился во владении и пользовании (собственником или кем-то другим) для нужд сельского хозяйства не менее 7 лет в течение 10 лет непосредственно перед смертью
Облегчение ограничено тем, что было бы доступно до замены.
Если имущество было продано по цене ниже рыночной, компенсация ограничивается той долей, которая была дана в качестве подарка.
Пример
Джон продает поле на своей ферме стоимостью 2000 фунтов стерлингов своему сыну Дэвиду на соседней ферме за 500 фунтов стерлингов. Это означает, что он дарит Дэвиду 75% стоимости поля = 1500 фунтов стерлингов.
Год спустя Дэвид продает поле и использует 2000 фунтов стерлингов от продажи, чтобы купить другое поле за 2000 фунтов стерлингов.
Джон умирает через 3 года после продажи.
При расчете IHT в качестве подарка, сделанного Джон и имеет право на сельскохозяйственную помощь.
Бизнес-помощь
Вы не можете требовать коммерческую помощь на стоимость актива, на который вы уже заявили сельскохозяйственную помощь. Тем не менее, фермерское предприятие может обнаружить, что Business Relief предоставляется на стоимость актива, не полностью покрываемого сельскохозяйственной помощью, если условия соблюдены.
Y-хромосома: вне определения пола
Y-хромосома: вне определения полаРозанна Ф. Чжао, доктор философии.
NIH MD/Ph.D. Партнерская учебная программа Scholar Геном человека состоит из 23 пар хромосом (22 пары аутосом и одна пара половых хромосом), причем каждый родитель вносит по одной хромосоме на пару. Хромосомы X и Y, также известные как половые хромосомы, определяют биологический пол человека: женщины наследуют X-хромосому от отца для генотипа XX, а мужчины наследуют Y-хромосому от отца для генотипа XY (только матери). проходят по Х-хромосомам).Наличие или отсутствие Y-хромосомы имеет решающее значение, поскольку она содержит гены, необходимые для преодоления биологического значения по умолчанию — женского развития — и обеспечения развития мужской репродуктивной системы.
Хотя роль Y-хромосомы в определении пола ясна, исследования показали, что она подвергается быстрому эволюционному износу. Много поколений назад Y-хромосома была большой и содержала столько же генов, сколько и X-хромосома. Теперь он в несколько раз меньше своего прошлого размера и содержит менее 80 функциональных генов.На протяжении многих лет это приводило к спорам и опасениям относительно возможной судьбы Y-хромосомы. Многие предполагают, что Y-хромосома стала лишней и может полностью распасться в течение следующих 10 миллионов лет. В то время как исследования Y-хромосомы были сложными из-за палиндромной и богатой повторами природы ее последовательности ДНК, недавние достижения в области геномики дали некоторые неожиданные результаты.
В этом выпуске геномного продвижения месяца освещаются два исследования, опубликованные в выпуске Nature от 24 апреля 2014 г., в которых исследуется эволюционный путь Y-хромосомы у различных млекопитающих.Вместе эти исследования демонстрируют стабильность Y-хромосомы за последние 25 миллионов лет. Они также раскрывают некоторые критические функции Y-хромосомы, которые предполагают, что она может остаться.
Для начала давайте углубимся в эволюционное происхождение половых хромосом примерно 200-300 миллионов лет назад. Хромосомы X и Y, происходящие из аутосом, изначально были примерно одинакового размера. В какой-то определенный момент Y-хромосома постепенно потеряла способность рекомбинировать — или обмениваться генетической информацией — с X-хромосомой и начала развиваться независимо.Это быстро привело к катастрофическому износу Y-хромосомы, которая теперь содержит только 3 процента генов, которые когда-то были общими с X-хромосомой.
Недавняя работа исследовательской группы Дэвида К. Пейджа, доктора медицины, из Института Уайтхеда, Массачусетского технологического института, и Хенрика Кессманна, доктора философии, из Швейцарского института биоинформатики и Лозаннского университета в Швейцарии, предполагает что первоначально быстрое снижение Y-хромосомы могло выровняться и стабилизироваться.
Используя разные геномные технологии, эти две исследовательские группы независимо проанализировали эволюцию Y-хромосомы у двух отдельных наборов млекопитающих, охватывающих более 15 различных видов, включая людей, шимпанзе, макак-резусов, быков, мартышек, мышей, крыс, собак. и опоссумы. Поразительно, но они обнаружили небольшую, но стабильную группу важных регуляторных генов на Y-хромосоме, которые сохранялись в течение длительного эволюционного периода, даже когда окружающие гены разрушались.Примечательно, что эти гены играют критически важную роль в управлении экспрессией других генов по всему геному и могут воздействовать на ткани по всему телу человека. Одна из причин длительной выносливости этих регуляторных генов Y-хромосомы заключается в том, что они «зависят от дозы», что означает, что для нормального функционирования требуются две копии.
Для большинства генов Х-хромосомы требуется только одна копия. У женщин есть две Х-хромосомы и, следовательно, две копии каждого Х-сцепленного гена, поэтому одна копия случайно инактивирована или выключена.У мужчин есть только одна Х-хромосома, поэтому экспрессируется только одна копия.
Однако регуляторные гены часто являются дозозависимыми и гапло-недостаточными, т. е. требуются две копии гена, а наличие только одной копии может привести к аномалиям или заболеванию. У самок эти регуляторные гены избегают Х-инактивации, так что копия на второй Х-хромосоме также экспрессируется; у мужчин, имеющих только одну Х-хромосому, сохранение этой группы регуляторных генов на Y-хромосоме имеет решающее значение для обеспечения второй копии.
В целом это означает, что помимо своей роли в определении пола и фертильности Y-хромосома также содержит важные гены, которые имеют решающее значение для здоровья и выживания мужчин.
Эти результаты имеют большое значение для нашего понимания различий в биологии, здоровье и заболеваниях между мужчинами и женщинами. Поскольку гены на X- и Y-хромосомах имеют историю независимого отбора друг от друга, могут существовать тонкие функциональные различия, являющиеся прямым следствием генетических различий на двух хромосомах.Хотя эти различия еще не были подробно изучены, дополнительные исследования консервативных генов Y-хромосомы могут помочь нам понять различия в базовой биологии и восприимчивости к заболеваниям у мужчин и женщин и лучше управлять здоровьем.
Читать статьи:
Беллотт Д.В., Хьюз Дж.Ф., Скалецкий Х., Браун Л.Г., Пынтикова Т., Чо Т.Дж., Куцева Н., Заглул С., Грейвс Т., Рок С., Кремицки С., Фултон Р.С., Дуган С., Дин Ю., Мортон Д., Хан З., Льюис Л.
 Поиск по сайту
Поиск по сайту